

What yeast and cardiomyocytes share: ultradian oscillatory redox mechanisms of cellular coherence and survival
- PMID: 22143867
- PMCID: PMC3348865
- DOI: 10.1039/c1ib00124h
What yeast and cardiomyocytes share: ultradian oscillatory redox mechanisms of cellular coherence and survival
Abstract
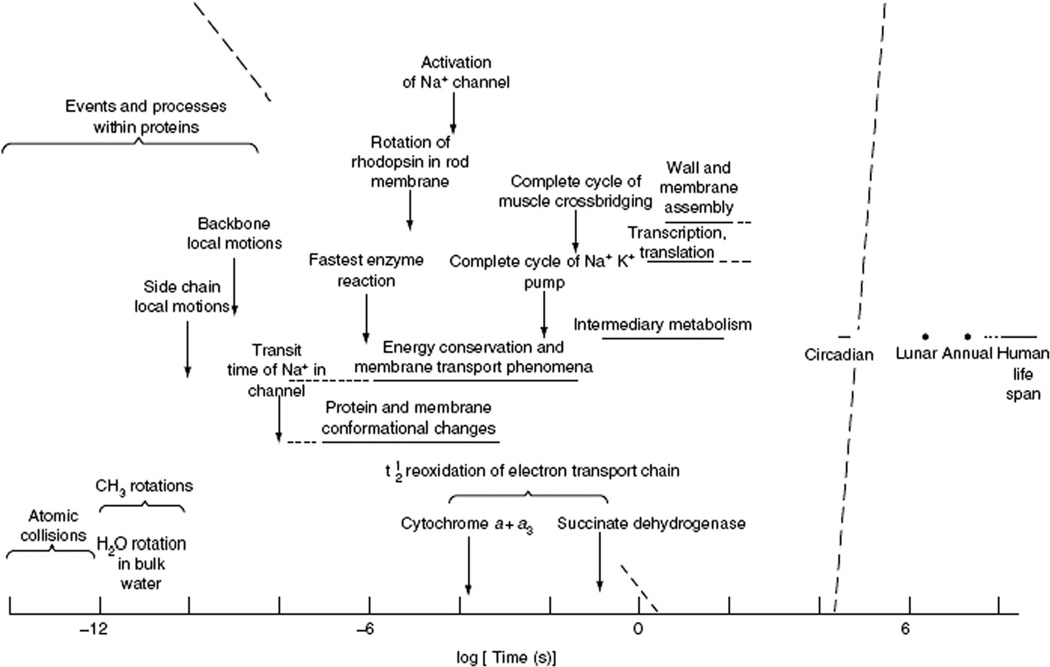
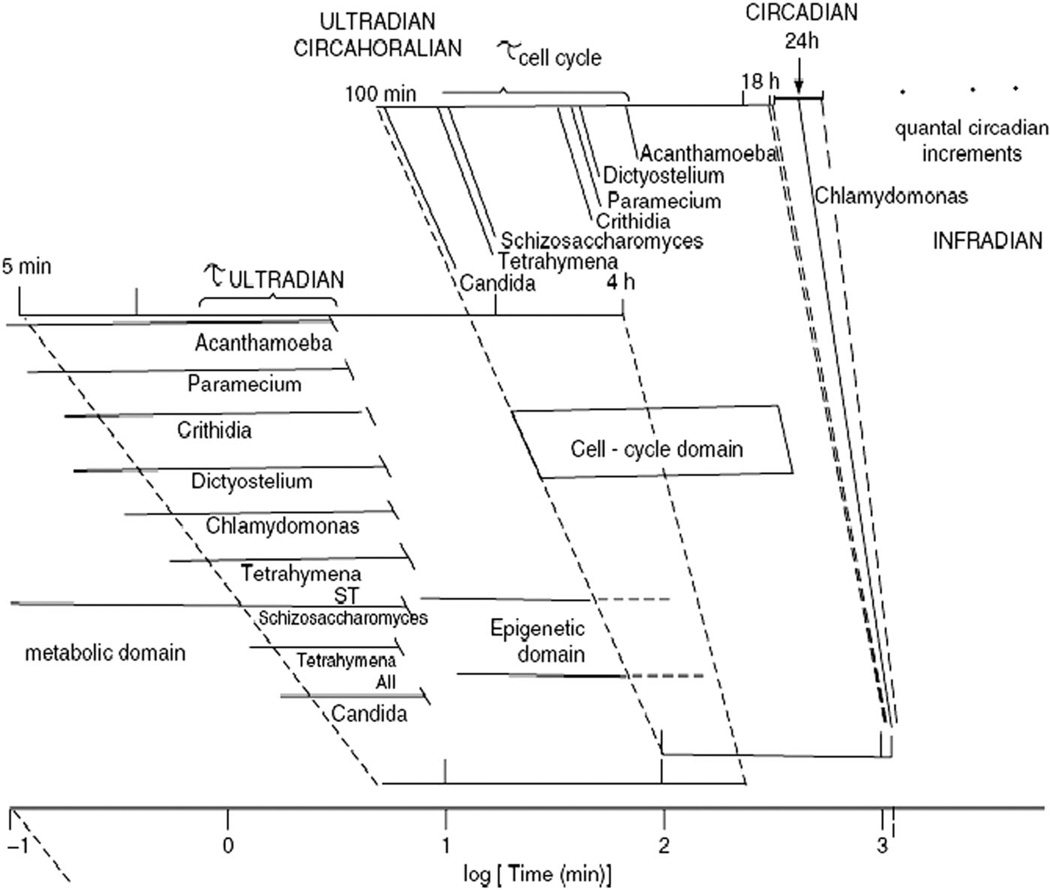
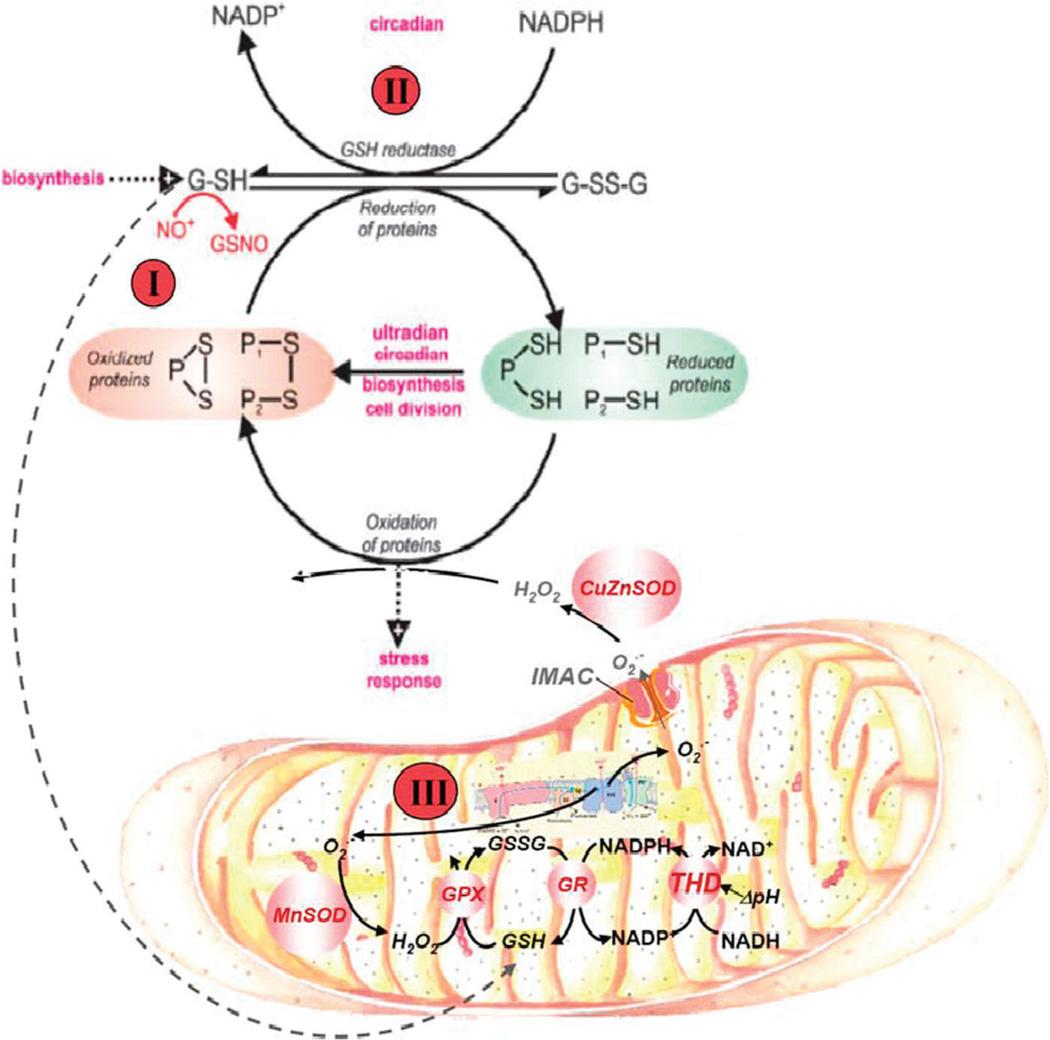
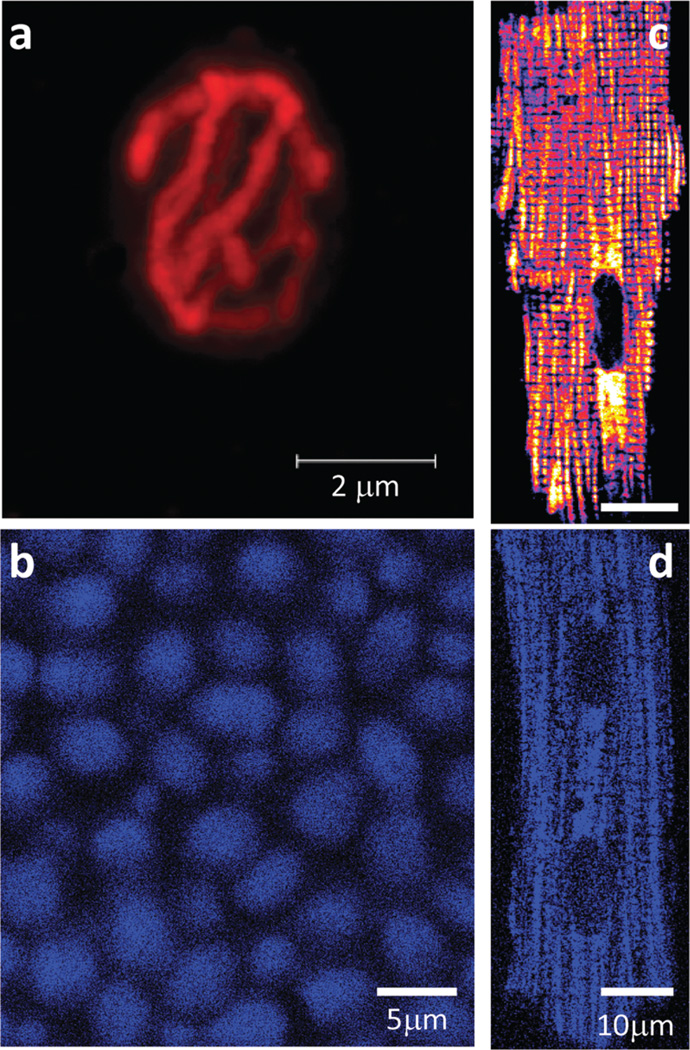
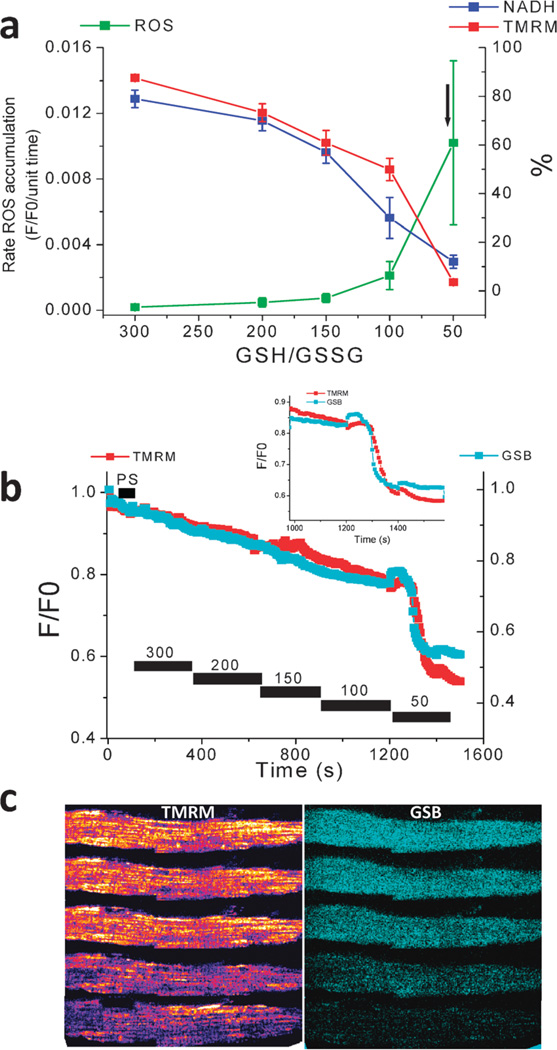
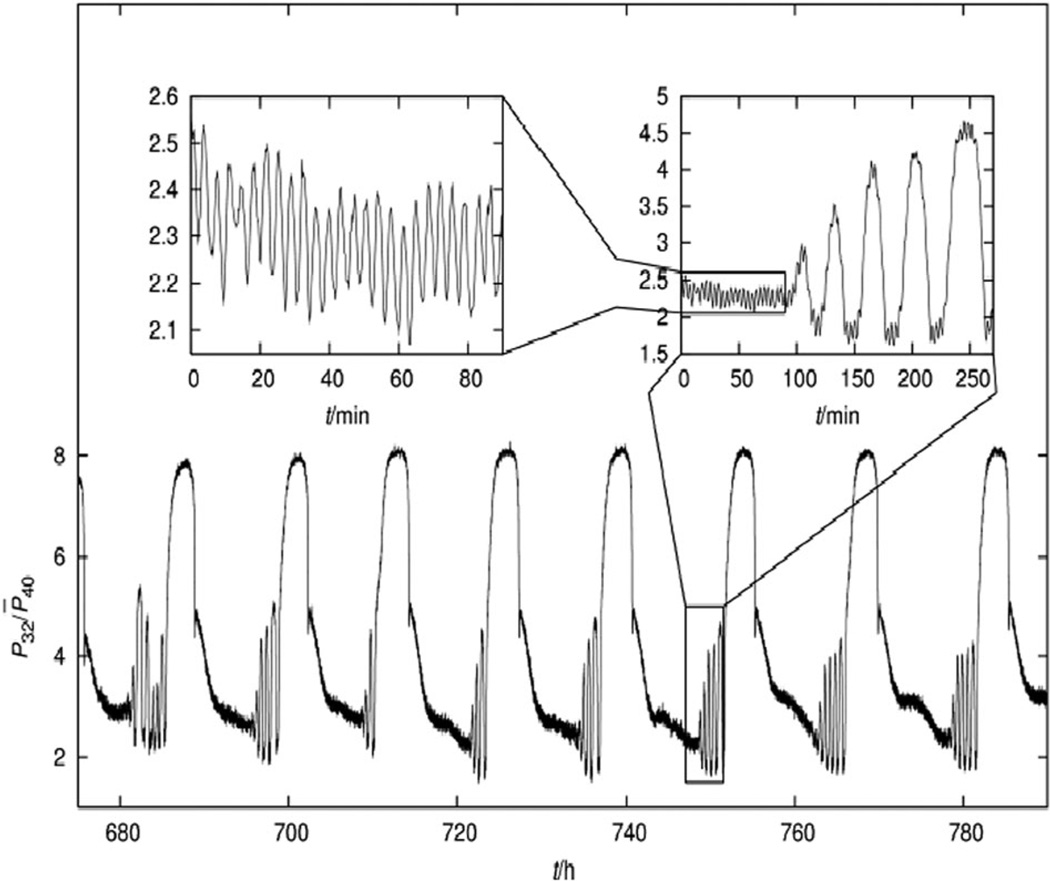
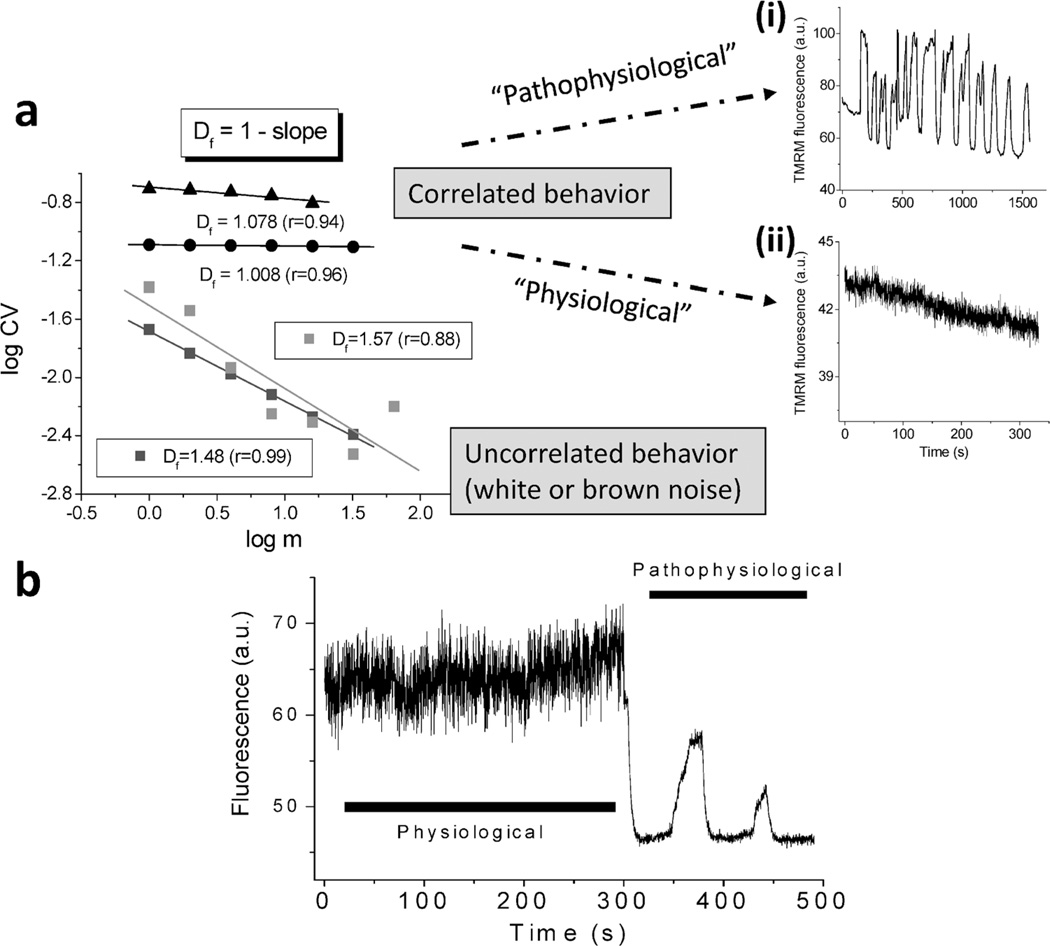
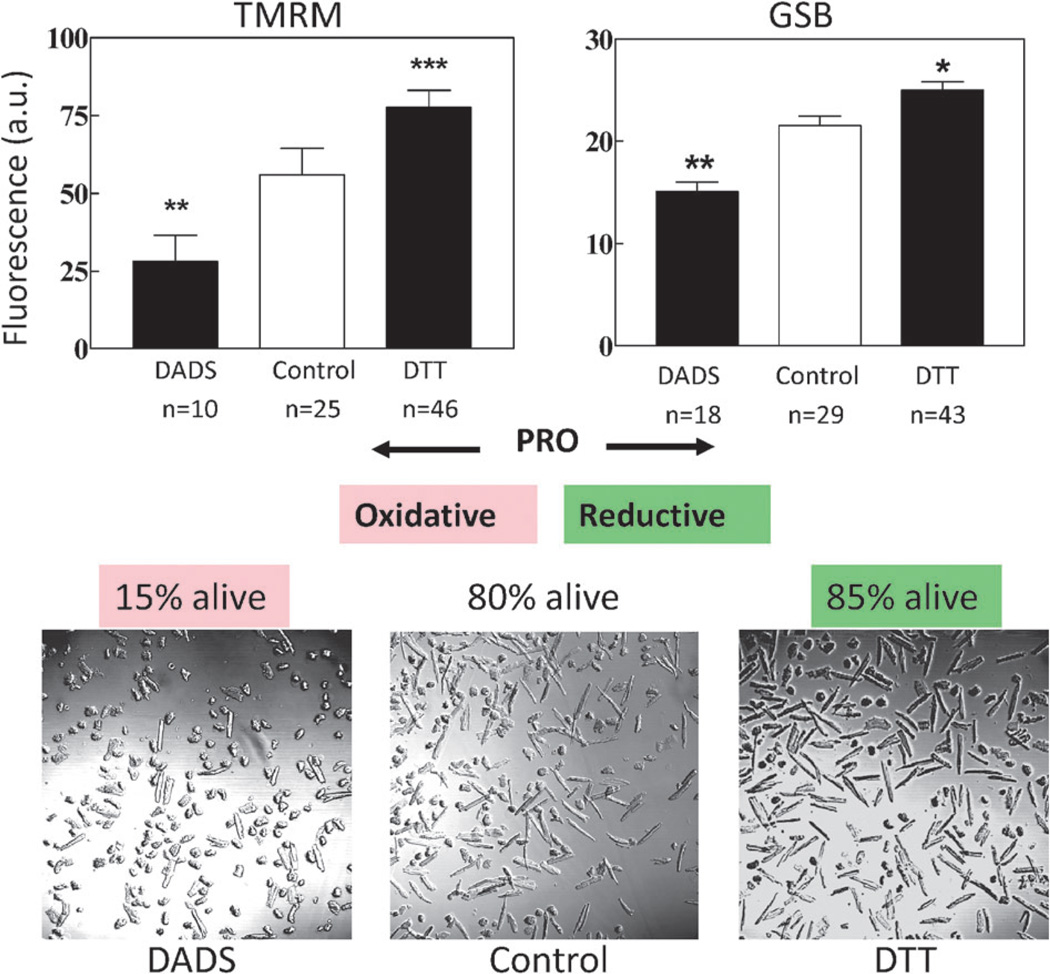
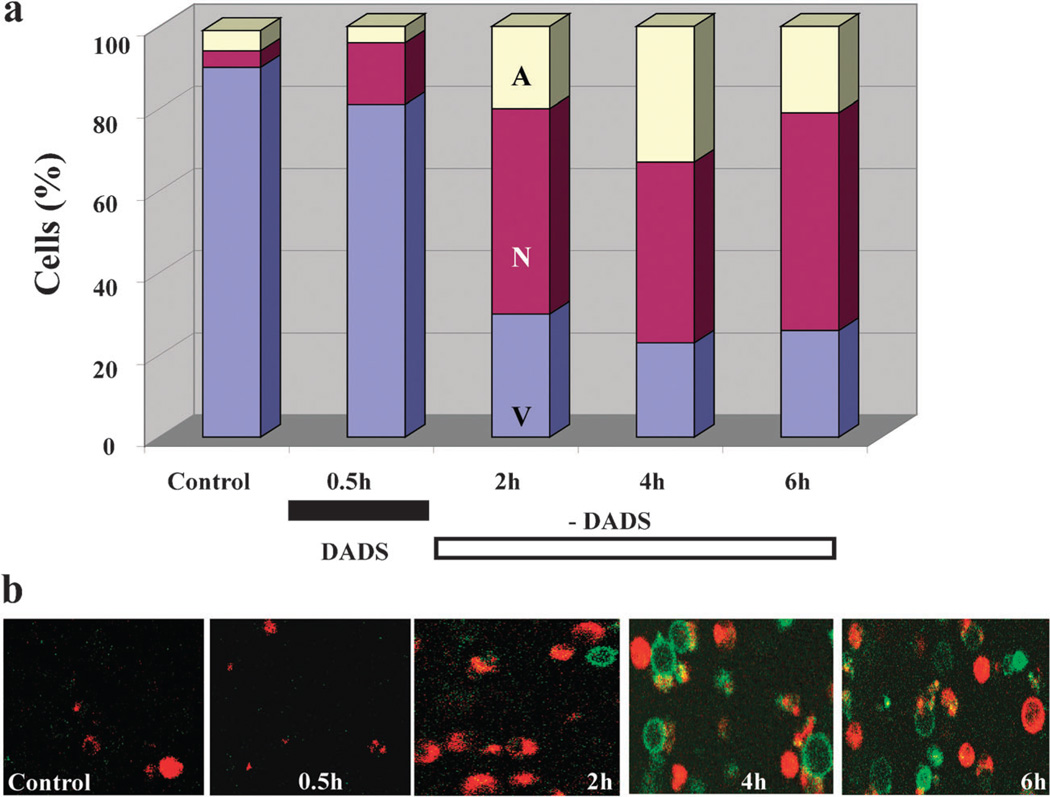
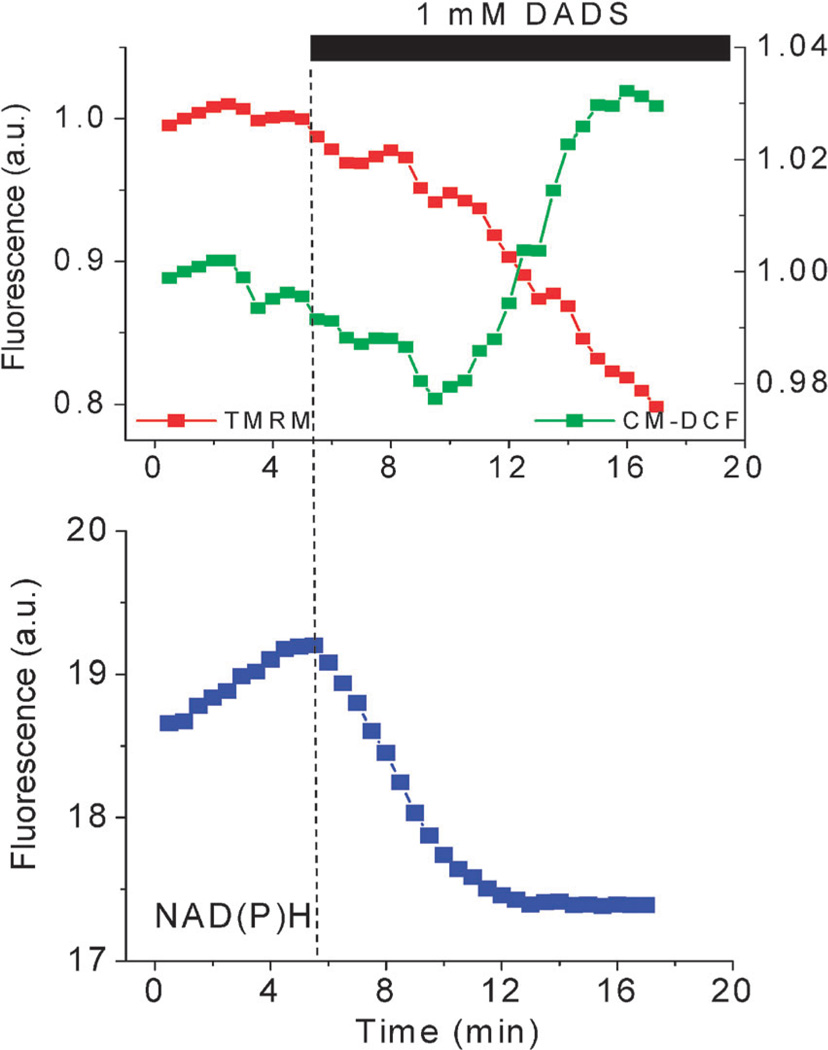
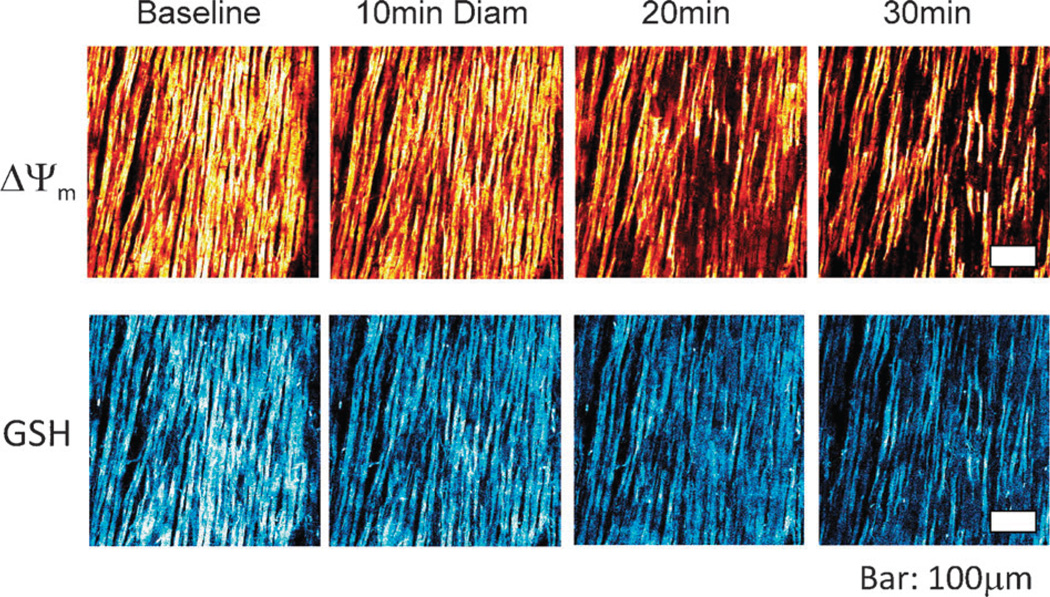
The coherent and robust, yet sensitively adaptable, nature of organisms is an astonishing phenomenon that involves massive parallel processing and concerted network performance at the molecular level. Unravelling the dynamic complexities of the living state underlines the essential operation of ultradian oscillations, rhythms and clocks for the establishment and maintenance of functional order simultaneously on fast and slower timescales. Non-invasive monitoring of respiration, mitochondrial inner membrane potentials, and redox states (especially those of NAD(P)H, flavin, and the monochlorobimane complex of glutathione), even after more than 50 years research, continue to provide both new insights and biomedical applications. Experiments with yeast and in cardiac cells reveal astonishing parallels and similarities in their dynamic biochemical organization.
This journal is © The Royal Society of Chemistry 2012
Figures
Similar articles
-
Respiratory oscillations in yeast: mitochondrial reactive oxygen species, apoptosis and time; a hypothesis.FEMS Yeast Res. 2003 Jun;3(4):333-9. doi: 10.1016/S1567-1356(03)00071-0. FEMS Yeast Res. 2003. PMID: 12748046 Review.
-
Respiratory oscillations in yeasts.Adv Exp Med Biol. 2008;641:118-40. doi: 10.1007/978-0-387-09794-7_9. Adv Exp Med Biol. 2008. PMID: 18783176 Review.
-
The scale-free dynamics of eukaryotic cells.PLoS One. 2008;3(11):e3624. doi: 10.1371/journal.pone.0003624. Epub 2008 Nov 4. PLoS One. 2008. PMID: 18982073 Free PMC article.
-
Ultradian metronome: timekeeper for orchestration of cellular coherence.Trends Biochem Sci. 2005 Jul;30(7):373-7. doi: 10.1016/j.tibs.2005.05.005. Trends Biochem Sci. 2005. PMID: 15935677 Review.
-
Cellular oscillators: rhythmic gene expression and metabolism.Curr Opin Cell Biol. 2005 Apr;17(2):223-9. doi: 10.1016/j.ceb.2005.01.007. Curr Opin Cell Biol. 2005. PMID: 15780601 Review.
Cited by
-
The fractal organization of ultradian rhythms in avian behavior.Sci Rep. 2017 Apr 6;7(1):684. doi: 10.1038/s41598-017-00743-2. Sci Rep. 2017. PMID: 28386121 Free PMC article.
-
Mitochondrial respiration and ROS emission during β-oxidation in the heart: An experimental-computational study.PLoS Comput Biol. 2017 Jun 9;13(6):e1005588. doi: 10.1371/journal.pcbi.1005588. eCollection 2017 Jun. PLoS Comput Biol. 2017. PMID: 28598967 Free PMC article.
-
Complex oscillatory redox dynamics with signaling potential at the edge between normal and pathological mitochondrial function.Front Physiol. 2014 Jul 8;5:257. doi: 10.3389/fphys.2014.00257. eCollection 2014. Front Physiol. 2014. PMID: 25071602 Free PMC article.
-
Mitochondrial redox and pH signaling occurs in axonal and synaptic organelle clusters.Sci Rep. 2016 Mar 22;6:23251. doi: 10.1038/srep23251. Sci Rep. 2016. PMID: 27000952 Free PMC article.
-
A dynamically coherent pattern of rhythms that matches between distant species across the evolutionary scale.Sci Rep. 2023 Apr 1;13(1):5326. doi: 10.1038/s41598-023-32286-0. Sci Rep. 2023. PMID: 37005423 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
