

Langerhans cell antigen capture through tight junctions confers preemptive immunity in experimental staphylococcal scalded skin syndrome
- PMID: 22143886
- PMCID: PMC3244045
- DOI: 10.1084/jem.20111718
Langerhans cell antigen capture through tight junctions confers preemptive immunity in experimental staphylococcal scalded skin syndrome
Abstract
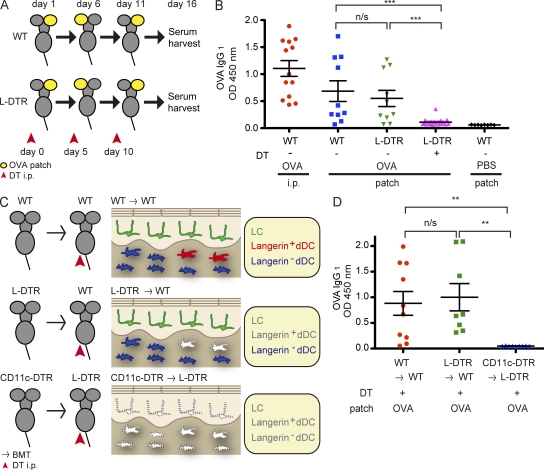
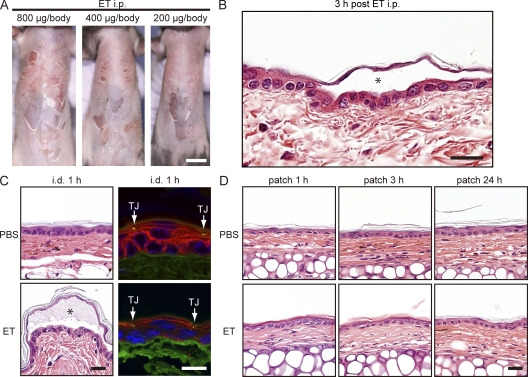
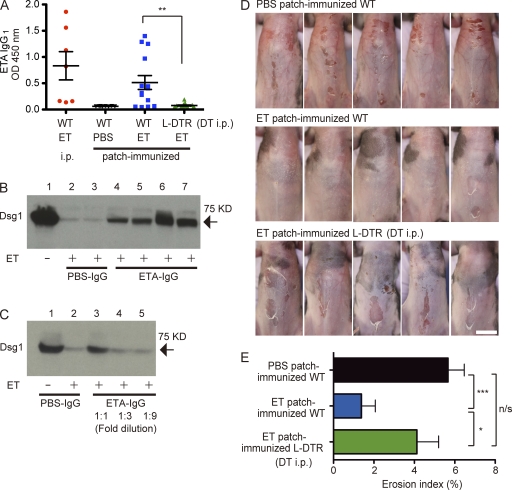
Epidermal Langerhans cells (LCs) extend dendrites through tight junctions (TJs) to survey the skin surface, but their immunological contribution in vivo remains elusive. We show that LCs were essential for inducing IgG(1) responses to patch-immunized ovalbumin in mice that lacked skin dendritic cell subsets. The significance of LC-induced humoral responses was demonstrated in a mouse model of staphylococcal scalded skin syndrome (SSSS), a severe blistering disease in which the desmosomal protein Dsg1 (desmoglein1) is cleaved by Staphylococcus aureus-derived exfoliative toxin (ET). Importantly, ET did not penetrate TJs, and patch immunization did not alter epidermal integrity. Nevertheless, neutralizing anti-ET IgG(1) was induced after patch immunization and abolished upon LC depletion, indicating that antigen capture through TJs by LCs induced humoral immunity. Strikingly, the ET-patched mice were protected from developing SSSS after intraperitoneal ET challenge, whereas LC-depleted mice were susceptible to SSSS, demonstrating a vital role for LC-induced IgG(1) in systemic defense against circulating toxin in vivo. Therefore, LCs elicit humoral immunity to antigens that have not yet violated the epidermal barrier, providing preemptive immunity against potentially pathogenic skin microbes. Targeting this immunological process confers protection with minimal invasiveness and should have a marked impact on future strategies for development of percutaneous vaccines.
Figures
Similar articles
-
Toxin levels in serum correlate with the development of staphylococcal scalded skin syndrome in a murine model.Infect Immun. 2001 Aug;69(8):5193-7. doi: 10.1128/IAI.69.8.5193-5197.2001. Infect Immun. 2001. PMID: 11447206 Free PMC article.
-
[Immunological investigations on pathogenesis of staphylococcal scalded skin syndrome].Rinsho Byori. 1995 Jun;43(6):547-56. Rinsho Byori. 1995. PMID: 7602798 Review. Japanese.
-
Staphylococcal scalded skin syndrome.J Med Microbiol. 1995 Nov;43(5):318-27. doi: 10.1099/00222615-43-5-318. J Med Microbiol. 1995. PMID: 7562995 Review.
-
Distinct behavior of human Langerhans cells and inflammatory dendritic epidermal cells at tight junctions in patients with atopic dermatitis.J Allergy Clin Immunol. 2014 Oct;134(4):856-64. doi: 10.1016/j.jaci.2014.08.001. J Allergy Clin Immunol. 2014. PMID: 25282566
-
Exfoliative toxin A staphylococcal scalded skin syndrome in preterm infants.Eur J Pediatr. 2015 Apr;174(4):551-5. doi: 10.1007/s00431-014-2414-3. Epub 2014 Sep 7. Eur J Pediatr. 2015. PMID: 25194957
Cited by
-
(Not) Home alone: Antigen presenting cell - T Cell communication in barrier tissues.Front Immunol. 2022 Sep 29;13:984356. doi: 10.3389/fimmu.2022.984356. eCollection 2022. Front Immunol. 2022. PMID: 36248804 Free PMC article. Review.
-
N-Linked Glycans Shape Skin Immune Responses during Arthritis and Myositis after Intradermal Infection with Ross River Virus.J Virol. 2022 Sep 14;96(17):e0099922. doi: 10.1128/jvi.00999-22. Epub 2022 Aug 24. J Virol. 2022. PMID: 36000846 Free PMC article.
-
Treatment-resistant atopic dermatitis: challenges and solutions.Clin Cosmet Investig Dermatol. 2019 Mar 21;12:181-192. doi: 10.2147/CCID.S163814. eCollection 2019. Clin Cosmet Investig Dermatol. 2019. PMID: 30962700 Free PMC article. Review.
-
One Size Does Not Fit All: Diversifying Immune Function in the Skin.J Immunol. 2022 Jan 15;208(2):227-234. doi: 10.4049/jimmunol.2100758. J Immunol. 2022. PMID: 35017212 Free PMC article. Review.
-
EpCAM Expressed by Murine Epidermal Langerhans Cells Modulates Immunization to an Epicutaneously Applied Protein Antigen.J Invest Dermatol. 2016 Aug;136(8):1627-1635. doi: 10.1016/j.jid.2016.04.005. Epub 2016 Apr 19. J Invest Dermatol. 2016. PMID: 27106675 Free PMC article.
References
-
- Bennett C.L., Noordegraaf M., Martina C.A., Clausen B.E. 2007. Langerhans cells are required for efficient presentation of topically applied hapten to T cells. J. Immunol. 179:6830–6835 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
