

Scrambling eggs: meiotic drive and the evolution of female recombination rates
- PMID: 22143919
- PMCID: PMC3276612
- DOI: 10.1534/genetics.111.136721
Scrambling eggs: meiotic drive and the evolution of female recombination rates
Abstract
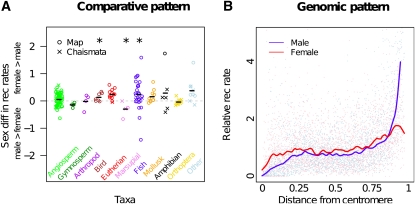
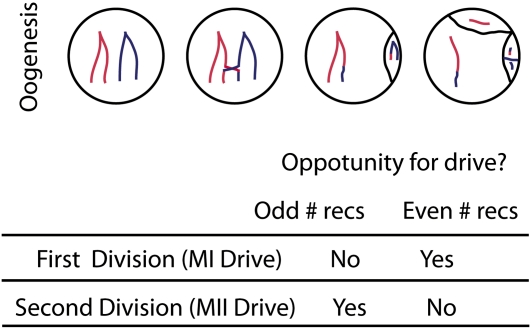
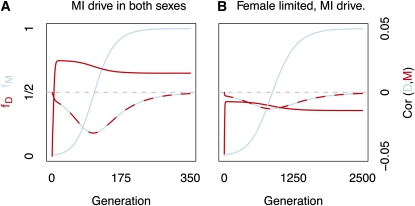
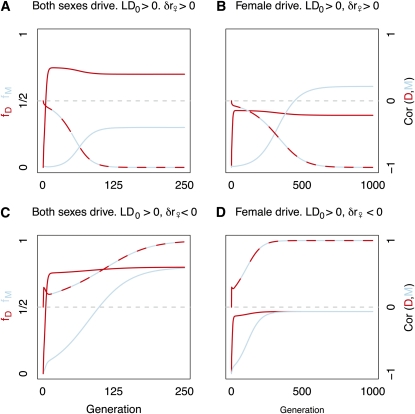
Theories to explain the prevalence of sex and recombination have long been a central theme of evolutionary biology. Yet despite decades of attention dedicated to the evolution of sex and recombination, the widespread pattern of sex differences in the recombination rate is not well understood and has received relatively little theoretical attention. Here, we argue that female meiotic drivers--alleles that increase in frequency by exploiting the asymmetric cell division of oogenesis--present a potent selective pressure favoring the modification of the female recombination rate. Because recombination plays a central role in shaping patterns of variation within and among dyads, modifiers of the female recombination rate can function as potent suppressors or enhancers of female meiotic drive. We show that when female recombination modifiers are unlinked to female drivers, recombination modifiers that suppress harmful female drive can spread. By contrast, a recombination modifier tightly linked to a driver can increase in frequency by enhancing female drive. Our results predict that rapidly evolving female recombination rates, particularly around centromeres, should be a common outcome of meiotic drive. We discuss how selection to modify the efficacy of meiotic drive may contribute to commonly observed patterns of sex differences in recombination.
Figures
References
-
- Aparicio J. M., Ortego J., Calabuig G., Cordero P. J., 2010. Evidence of subtle departures from Mendelian segregation in a wild lesser kestrel (Falco naumanni) population. Heredity 105(2): 213–219 - PubMed
-
- Axelsson E., Albrechtsen A., Van A. P., Li L., Megens H. J., et al. , 2010. Segregation distortion in chicken and the evolutionary consequences of female meiotic drive in birds. Heredity 105(3): 290–298 - PubMed
-
- Barton N. H., 2009. Why sex and recombination? Cold Spring Harbor Symp. Quant. Biol. 74(5385): 187–195 - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
