

Sialylation of lipooligosaccharides is dispensable for the virulence of Haemophilus ducreyi in humans
- PMID: 22144477
- PMCID: PMC3264291
- DOI: 10.1128/IAI.05826-11
Sialylation of lipooligosaccharides is dispensable for the virulence of Haemophilus ducreyi in humans
Abstract
Sialylated glycoconjugates on the surfaces of mammalian cells play important roles in intercellular communication and self-recognition. The sialic acid preferentially expressed in human tissues is N-acetylneuraminic acid (Neu5Ac). In a process called molecular mimicry, many bacterial pathogens decorate their cell surface glycolipids with Neu5Ac. Incorporation of Neu5Ac into bacterial glycolipids promotes bacterial interactions with host cell receptors called Siglecs. These interactions affect bacterial adherence, resistance to serum killing and phagocytosis, and innate immune responses. Haemophilus ducreyi, the etiologic agent of chancroid, expresses lipooligosaccharides (LOS) that are highly sialylated. However, an H. ducreyi sialyltransferase (lst) mutant, whose LOS contain reduced levels of Neu5Ac, is fully virulent in human volunteers. Recently, a second sialyltransferase gene (Hd0053) was discovered in H. ducreyi, raising the possibility that Hd0053 compensated for the loss of lst during human infection. CMP-Neu5Ac is the obligate nucleotide sugar donor for all bacterial sialyltransferases; LOS derived from an H. ducreyi CMP-Neu5Ac synthetase (neuA) mutant has no detectable Neu5Ac. Here, we compared an H. ducreyi neuA mutant to its wild-type parent in several models of pathogenesis. In human inoculation experiments, the neuA mutant formed papules and pustules at rates that were no different than those of its parent. When grown in media with and without Neu5Ac supplementation, the neuA mutant and its parent had similar phenotypes in bactericidal, macrophage uptake, and dendritic cell activation assays. Although we cannot preclude a contribution of LOS sialylation to ulcerative disease, these data strongly suggest that sialylation of LOS is dispensable for H. ducreyi pathogenesis in humans.
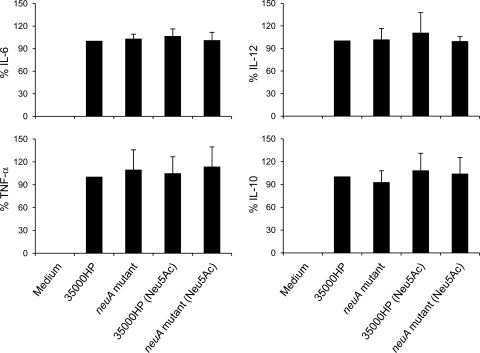
Figures
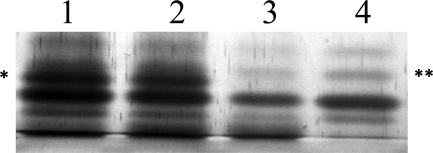
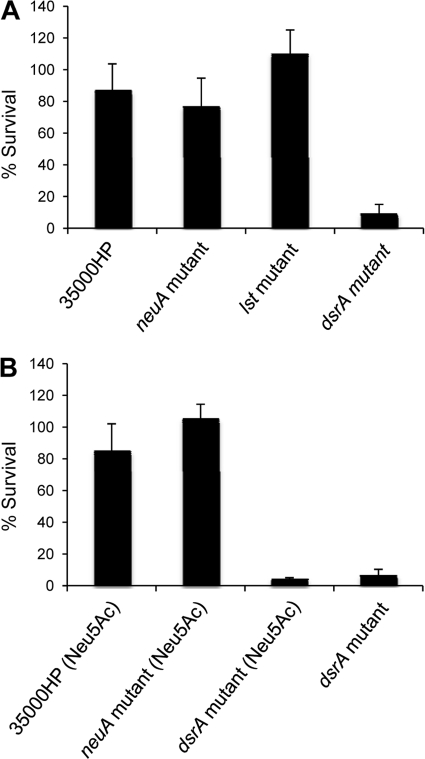
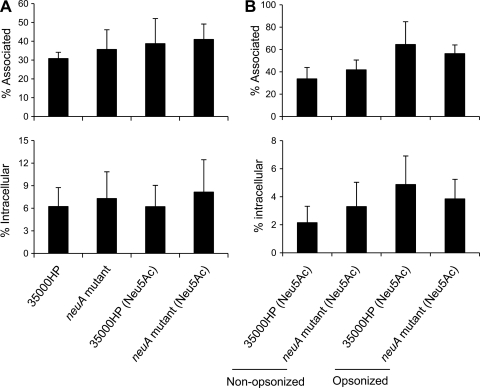
Similar articles
-
Haemophilus ducreyi produces a novel sialyltransferase. Identification of the sialyltransferase gene and construction of mutants deficient in the production of the sialic acid-containing glycoform of the lipooligosaccharide.J Biol Chem. 1999 Feb 12;274(7):4106-14. doi: 10.1074/jbc.274.7.4106. J Biol Chem. 1999. PMID: 9933604
-
The Hd0053 gene of Haemophilus ducreyi encodes an alpha2,3-sialyltransferase.Biochem Biophys Res Commun. 2007 Sep 21;361(2):555-60. doi: 10.1016/j.bbrc.2007.07.063. Epub 2007 Jul 23. Biochem Biophys Res Commun. 2007. PMID: 17662691 Free PMC article.
-
Expression of sialylated or paragloboside-like lipooligosaccharides are not required for pustule formation by Haemophilus ducreyi in human volunteers.Infect Immun. 1999 Dec;67(12):6335-40. doi: 10.1128/IAI.67.12.6335-6340.1999. Infect Immun. 1999. PMID: 10569746 Free PMC article.
-
Interactions of the Skin Pathogen Haemophilus ducreyi With the Human Host.Front Immunol. 2021 Feb 3;11:615402. doi: 10.3389/fimmu.2020.615402. eCollection 2020. Front Immunol. 2021. PMID: 33613541 Free PMC article. Review.
-
Host-pathogen interplay of Haemophilus ducreyi.Curr Opin Infect Dis. 2010 Feb;23(1):64-9. doi: 10.1097/QCO.0b013e328334c0cb. Curr Opin Infect Dis. 2010. PMID: 19918177 Free PMC article. Review.
Cited by
-
Biological functions of sialic acid as a component of bacterial endotoxin.Front Microbiol. 2022 Oct 20;13:1028796. doi: 10.3389/fmicb.2022.1028796. eCollection 2022. Front Microbiol. 2022. PMID: 36338080 Free PMC article. Review.
-
Salmonella O48 Serum Resistance is Connected with the Elongation of the Lipopolysaccharide O-Antigen Containing Sialic Acid.Int J Mol Sci. 2017 Sep 21;18(10):2022. doi: 10.3390/ijms18102022. Int J Mol Sci. 2017. PMID: 28934165 Free PMC article.
-
Activation of CpxRA in Haemophilus ducreyi primarily inhibits the expression of its targets, including major virulence determinants.J Bacteriol. 2013 Aug;195(15):3486-502. doi: 10.1128/JB.00372-13. Epub 2013 May 31. J Bacteriol. 2013. PMID: 23729647 Free PMC article.
-
Sweet impersonators: Molecular mimicry of host glycans by bacteria.Glycobiology. 2022 Feb 26;32(1):11-22. doi: 10.1093/glycob/cwab104. Glycobiology. 2022. PMID: 34939094 Free PMC article. Review.
-
The Haemophilus ducreyi Fis protein is involved in controlling expression of the lspB-lspA2 operon and other virulence factors.Infect Immun. 2013 Nov;81(11):4160-70. doi: 10.1128/IAI.00714-13. Epub 2013 Aug 26. Infect Immun. 2013. PMID: 23980107 Free PMC article.
References
-
- Alfa MJ, Degagne P. 1997. Attachment of Haemophilus ducreyi to human foreskin fibroblasts involves LOS and fibronectin. Microb. Pathog. 22:39–46 - PubMed
-
- Al-Tawfiq JA, et al. 1998. Standardization of the experimental model of Haemophilus ducreyi infection in human subjects. J. Infect. Dis. 178:1684–1687 - PubMed
-
- Apicella MA, Griffiss JM, Schneider H. 1994. Isolation and characterization of lipopolysaccharides, lipooligosaccharides, and lipid A. Methods Enzymol. 235:242–252 - PubMed
-
- Apicella MA, et al. 1990. Modification by sialic acid of Neisseria gonorrhoeae lipooligosaccharide epitope expression in human urethral exudates: an immunoelectron microscopic analysis. J. Infect. Dis. 162:506–512 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
