

Role of CD11b/CD18 in the process of intoxication by the adenylate cyclase toxin of Bordetella pertussis
- PMID: 22144488
- PMCID: PMC3264305
- DOI: 10.1128/IAI.05979-11
Role of CD11b/CD18 in the process of intoxication by the adenylate cyclase toxin of Bordetella pertussis
Abstract
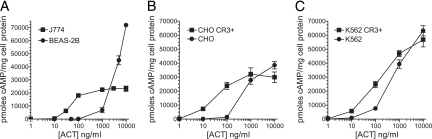
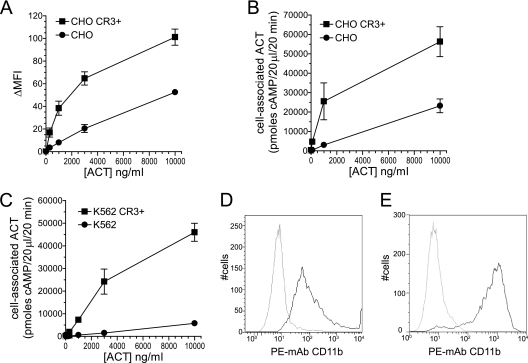
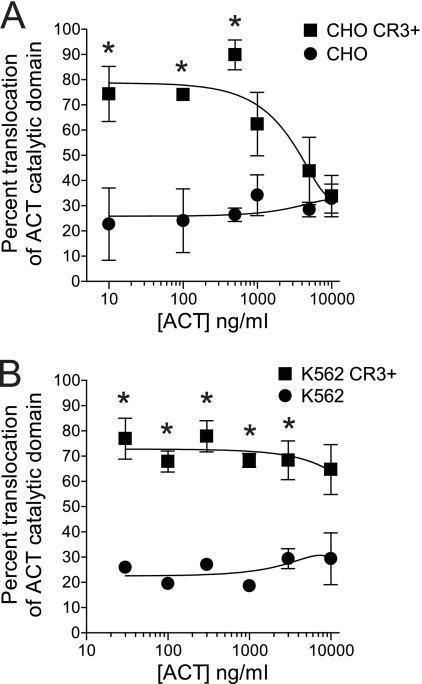
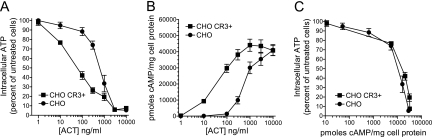
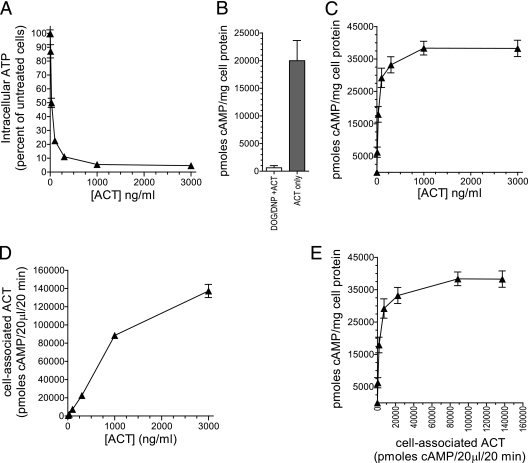
The adenylate cyclase toxin (ACT) of Bordetella pertussis does not require a receptor to generate intracellular cyclic AMP (cAMP) in a broad range of cell types. To intoxicate cells, ACT binds to the cell surface, translocates its catalytic domain across the cell membrane, and converts intracellular ATP to cAMP. In cells that express the integrin CD11b/CD18 (CR3), ACT is more potent than in CR3-negative cells. We find, however, that the maximum levels of cAMP accumulation inside CR3-positive and -negative cells are comparable. To better understand how CR3 affects the generation of cAMP, we used Chinese hamster ovary and K562 cells transfected to express CR3 and examined the steps in intoxication in the presence and absence of the integrin. The binding of ACT to cells is greater in CR3-expressing cells at all concentrations of ACT, and translocation of the catalytic domain is enhanced by CR3 expression, with ∼80% of ACT molecules translocating their catalytic domain in CR3-positive cells but only 25% in CR3-negative cells. Once in the cytosol, the unregulated catalytic domain converts ATP to cAMP, and at ACT concentrations >1,000 ng/ml, the intracellular ATP concentration is <5% of that in untreated cells, regardless of CR3 expression. This depletion of ATP prevents further production of cAMP, despite the CR3-mediated enhancement of binding and translocation. In addition to characterizing the effects of CR3 on the actions of ACT, these data show that ATP consumption is yet another concentration-dependent activity of ACT that must be considered when studying how ACT affects target cells.
Figures

Similar articles
-
Transmembrane segments of complement receptor 3 do not participate in cytotoxic activities but determine receptor structure required for action of Bordetella adenylate cyclase toxin.Pathog Dis. 2016 Apr;74(3):ftw008. doi: 10.1093/femspd/ftw008. Epub 2016 Jan 21. Pathog Dis. 2016. PMID: 26802078
-
Cyclic AMP-Elevating Capacity of Adenylate Cyclase Toxin-Hemolysin Is Sufficient for Lung Infection but Not for Full Virulence of Bordetella pertussis.Infect Immun. 2017 May 23;85(6):e00937-16. doi: 10.1128/IAI.00937-16. Print 2017 Jun. Infect Immun. 2017. PMID: 28396322 Free PMC article.
-
The adenylate cyclase toxin of Bordetella pertussis binds to target cells via the alpha(M)beta(2) integrin (CD11b/CD18).J Exp Med. 2001 May 7;193(9):1035-44. doi: 10.1084/jem.193.9.1035. J Exp Med. 2001. PMID: 11342588 Free PMC article.
-
Bordetella adenylate cyclase toxin: a unique combination of a pore-forming moiety with a cell-invading adenylate cyclase enzyme.Pathog Dis. 2015 Nov;73(8):ftv075. doi: 10.1093/femspd/ftv075. Epub 2015 Sep 20. Pathog Dis. 2015. PMID: 26391732 Free PMC article. Review.
-
Understanding the Mechanism of Translocation of Adenylate Cyclase Toxin across Biological Membranes.Toxins (Basel). 2017 Sep 21;9(10):295. doi: 10.3390/toxins9100295. Toxins (Basel). 2017. PMID: 28934133 Free PMC article. Review.
Cited by
-
Bordetella pertussis Adenylate Cyclase Toxin Disrupts Functional Integrity of Bronchial Epithelial Layers.Infect Immun. 2018 Feb 20;86(3):e00445-17. doi: 10.1128/IAI.00445-17. Print 2018 Mar. Infect Immun. 2018. PMID: 29203545 Free PMC article.
-
Nuanced differences in adenylate cyclase toxin production, acylation, and secretion may contribute to the evolution of virulence in Bordetella species.mBio. 2025 Jun 11;16(6):e0108225. doi: 10.1128/mbio.01082-25. Epub 2025 May 19. mBio. 2025. PMID: 40387377 Free PMC article.
-
Bordetella pertussis adenylate cyclase toxin translocation across a tethered lipid bilayer.Proc Natl Acad Sci U S A. 2013 Dec 17;110(51):20473-8. doi: 10.1073/pnas.1312975110. Epub 2013 Dec 2. Proc Natl Acad Sci U S A. 2013. PMID: 24297899 Free PMC article.
-
Rediscovering Pertussis.Front Pediatr. 2016 Jun 8;4:52. doi: 10.3389/fped.2016.00052. eCollection 2016. Front Pediatr. 2016. PMID: 27376050 Free PMC article. Review.
-
Phospholipase A activity of adenylate cyclase toxin mediates translocation of its adenylate cyclase domain.Proc Natl Acad Sci U S A. 2017 Aug 15;114(33):E6784-E6793. doi: 10.1073/pnas.1701783114. Epub 2017 Jul 31. Proc Natl Acad Sci U S A. 2017. PMID: 28760979 Free PMC article.
References
-
- Bachelet M, Richard MJ, Francois D, Polla BS. 2002. Mitochondrial alterations precede Bordetella pertussis-induced apoptosis. FEMS Immunol. Med. Microbiol. 32:125–131 - PubMed
-
- Basler M, et al. 2007. Segments crucial for membrane translocation and pore-forming activity of Bordetella adenylate cyclase toxin. J. Biol. Chem. 282:12419–12429 - PubMed
-
- Benz R, Maier E, Ladant D, Ullmann A, Sebo P. 1994. Adenylate-cyclase toxin (CyaA) of Bordetella pertussis. Evidence for the formation of small ion-permeable channels and comparison with HlyA of Escherichia coli. J. Biol. Chem. 269:27231–27239 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
