

Repetitive activity slows axonal conduction velocity and concomitantly increases mechanical activation threshold in single axons of the rat cranial dura
- PMID: 22144575
- PMCID: PMC3381306
- DOI: 10.1113/jphysiol.2011.220624
Repetitive activity slows axonal conduction velocity and concomitantly increases mechanical activation threshold in single axons of the rat cranial dura
Abstract
The passage of an action potential along a peripheral axon modulates the conduction velocity of subsequent action potentials. In C-neurones with unmyelinated axons repetitive activity progressively slows axonal conduction velocity and in microneurographic recordings from healthy human subjects the magnitude of this slowing can be used to predict the receptive properties of individual axons. Recently, a reduction in the number of available voltage-gated sodium channels (Na(V)) through inactivation has been implicated as the predominant factor responsible for the slowing of axonal conduction. Since Na(V)s are also responsible for the initiation of action potentials in sensory nerve terminals, changes in their availability may be expected to affect activation threshold for sensory stimuli. To examine this proposal, dynamic mechanical stimuli were used to make precise estimates of activation threshold in single unmyelinated axons innervating the rat cranial dura mater. Decreases in axonal conduction velocity induced by repetitive electrical stimulation were paralleled by an increase in mechanical activation threshold. Application of TTX (10-20 nM) also slowed axonal conduction velocity in all 11 fibres examined and in 9 of these this resulted in a parallel increase in mechanical activation threshold. We interpret this as indicating that a reduction in available Na(V) number contributes to both axonal conduction velocity slowing and the observed parallel increase in mechanical activation threshold. The slowing of axonal conduction velocity observed during repetitive activity thus represents a form of accommodation, i.e. self inhibition, which is likely to be decisive in limiting peripheral input to the spinal dorsal horn and thereby regulating processes that could otherwise lead to central sensitization.
Figures
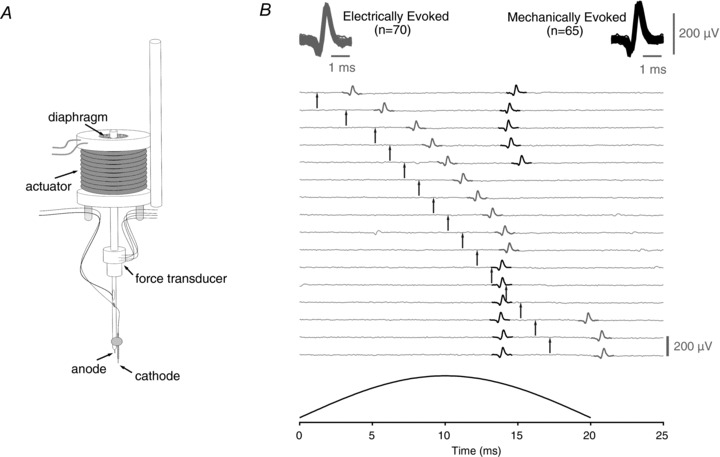
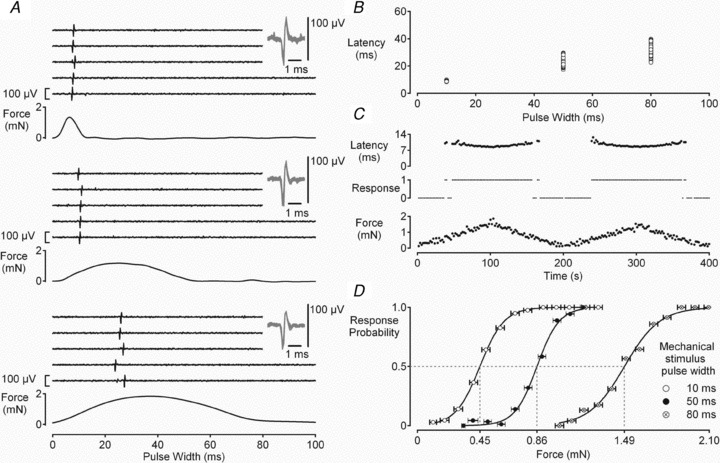
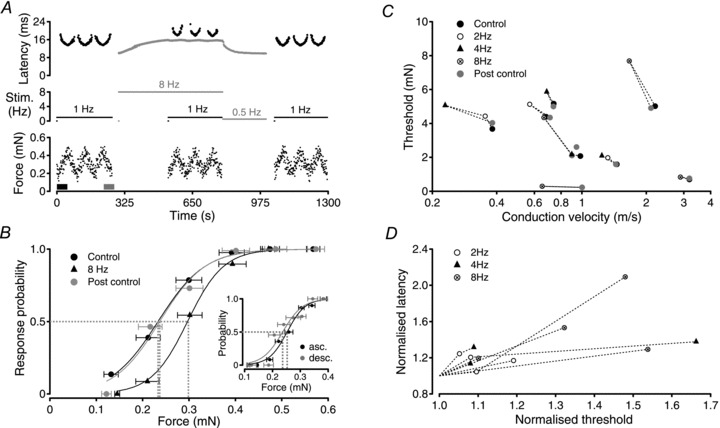
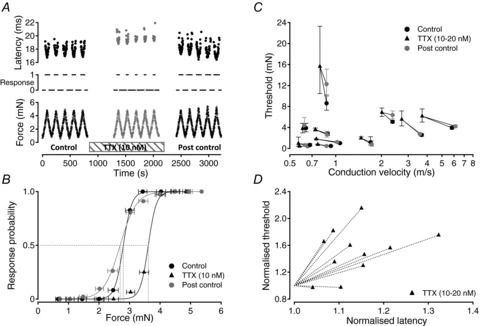
Comment in
-
Sodium channel slow inactivation and adaptation in C-fibres.J Physiol. 2012 Apr 1;590(7):1513-4. doi: 10.1113/jphysiol.2011.224436. J Physiol. 2012. PMID: 22467918 Free PMC article. No abstract available.
References
-
- Abrahamsen B, Zhao J, Asante CO, Cendan CM, Marsh S, Martinez-Barbera JP, Nassar MA, Dickenson AH, Wood JN. The cell and molecular basis of mechanical, cold, and inflammatory pain. Science. 2008;321:702–705. - PubMed
-
- Akopian AN, Sivilotti L, Wood JN. A tetrodotoxin-resistant voltage-gated sodium channel expressed by sensory neurons. Nature. 1996;379:257–262. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources