

Suppression of Gαs synthesis by simvastatin treatment of vascular endothelial cells
- PMID: 22144680
- PMCID: PMC3268423
- DOI: 10.1074/jbc.M111.303594
Suppression of Gαs synthesis by simvastatin treatment of vascular endothelial cells
Abstract
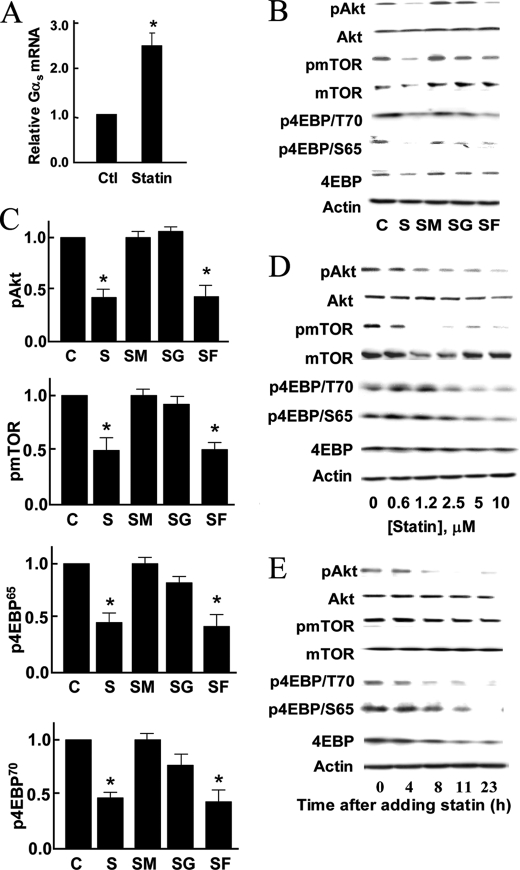
These studies explore the effects of statins on cyclic AMP-modulated signaling pathways in vascular endothelial cells. We previously observed (Kou, R., Sartoretto, J., and Michel, T. (2009) J. Biol. Chem. 284, 14734-14743) that simvastatin treatment of endothelial cells leads to a marked decrease in PKA-modulated phosphorylation of the protein VASP. Here we show that long-term treatment of mice with simvastatin attenuates the vasorelaxation response to the β-adrenergic agonist isoproterenol, without affecting endothelin-induced vasoconstriction or carbachol-induced vasorelaxation. We found that statin treatment of endothelial cells dose-dependently inhibits PKA activation as assessed by analyses of serine 157 VASP phosphorylation as well as Epac-mediated Rap1 activation. These effects of simvastatin are completely reversed by mevalonate and by geranylgeranyl pyrophosphate, implicating geranylgeranylation as a critical determinant of the stain response. We used biochemical approaches as well as fluorescence resonance energy transfer (FRET) methods with a cAMP biosensor to show that simvastatin treatment of endothelial cells markedly inhibits cAMP accumulation in response to epinephrine. Importantly, simvastatin treatment significantly decreases Gα(s) abundance, without affecting other Gα subunits. Simvastatin treatment does not influence Gα(s) protein stability, and paradoxically increases the abundance of Gα(s) mRNA. Finally, we found that simvastatin treatment inhibits Gα(s) translation mediated by Akt/mTOR/eIF4/4EBP. Taken together, these findings establish a novel mechanism by which simvastatin modulates β-adrenergic signaling in vascular wall, and may have implications for cardiovascular therapeutics.
Figures









Similar articles
-
Simvastatin suppresses leptin expression in 3T3-L1 adipocytes via activation of the cyclic AMP-PKA pathway induced by inhibition of protein prenylation.J Biochem. 2009 Jun;145(6):771-81. doi: 10.1093/jb/mvp035. Epub 2009 Mar 2. J Biochem. 2009. PMID: 19254925
-
Regulation of Rac1 by simvastatin in endothelial cells: differential roles of AMP-activated protein kinase and calmodulin-dependent kinase kinase-beta.J Biol Chem. 2009 May 29;284(22):14734-43. doi: 10.1074/jbc.M808664200. Epub 2009 Mar 30. J Biol Chem. 2009. PMID: 19332549 Free PMC article.
-
3-hydroxy-3-methylglutaryl coenzyme A reductase inhibitors interfere with angiogenesis by inhibiting the geranylgeranylation of RhoA.Circ Res. 2002 Jul 26;91(2):143-50. doi: 10.1161/01.res.0000028149.15986.4c. Circ Res. 2002. PMID: 12142347
-
Simvastatin modulates chemokine and chemokine receptor expression by geranylgeranyl isoprenoid pathway in human endothelial cells and macrophages.Atherosclerosis. 2006 Sep;188(1):51-8. doi: 10.1016/j.atherosclerosis.2005.10.015. Atherosclerosis. 2006. PMID: 16321392
-
Statin prevents tissue factor expression in human endothelial cells: role of Rho/Rho-kinase and Akt pathways.Circulation. 2002 Apr 16;105(15):1756-9. doi: 10.1161/01.cir.0000015465.73933.3b. Circulation. 2002. PMID: 11956113
Cited by
-
Metabolism-based approaches for autosomal dominant polycystic kidney disease.Front Mol Biosci. 2023 Feb 16;10:1126055. doi: 10.3389/fmolb.2023.1126055. eCollection 2023. Front Mol Biosci. 2023. PMID: 36876046 Free PMC article. Review.
-
Erythrocyte-derived microvesicles induce arterial spasms in JAK2V617F myeloproliferative neoplasm.J Clin Invest. 2020 May 1;130(5):2630-2643. doi: 10.1172/JCI124566. J Clin Invest. 2020. PMID: 32045382 Free PMC article.
-
Pravastatin-induced changes in expression of long non-coding and coding RNAs in endothelial cells.Physiol Rep. 2021 Jan;9(1):e14661. doi: 10.14814/phy2.14661. Physiol Rep. 2021. PMID: 33369888 Free PMC article.
-
Enhancement of autophagy by simvastatin through inhibition of Rac1-mTOR signaling pathway in coronary arterial myocytes.Cell Physiol Biochem. 2013;31(6):925-37. doi: 10.1159/000350111. Epub 2013 Jun 18. Cell Physiol Biochem. 2013. PMID: 23817226 Free PMC article.
-
Intracellular cAMP Sensor EPAC: Physiology, Pathophysiology, and Therapeutics Development.Physiol Rev. 2018 Apr 1;98(2):919-1053. doi: 10.1152/physrev.00025.2017. Physiol Rev. 2018. PMID: 29537337 Free PMC article. Review.
References
-
- Jain M. K., Ridker P. M. (2005) Anti-inflammatory effects of statins: clinical evidence and basic mechanisms. Nat. Rev. Drug Discov. 4, 977–987 - PubMed
-
- Mühlhäuser U., Zolk O., Rau T., Münzel F., Wieland T., Eschenhagen T. (2006) Atorvastatin desensitizes β-adrenergic signaling in cardiac myocytes via reduced isoprenylation of G-protein γ-subunits. FASEB J. 20, 785–787 - PubMed
-
- Igarashi J., Bernier S. G., Michel T. (2001) Sphingosine 1-phosphate and activation of endothelial nitric-oxide synthase. differential regulation of Akt and MAP kinase pathways by EDG and bradykinin receptors in vascular endothelial cells. J. Biol. Chem. 276, 12420–12426 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
