

Feeding cells induced by phytoparasitic nematodes require γ-tubulin ring complex for microtubule reorganization
- PMID: 22144887
- PMCID: PMC3228788
- DOI: 10.1371/journal.ppat.1002343
Feeding cells induced by phytoparasitic nematodes require γ-tubulin ring complex for microtubule reorganization
Abstract
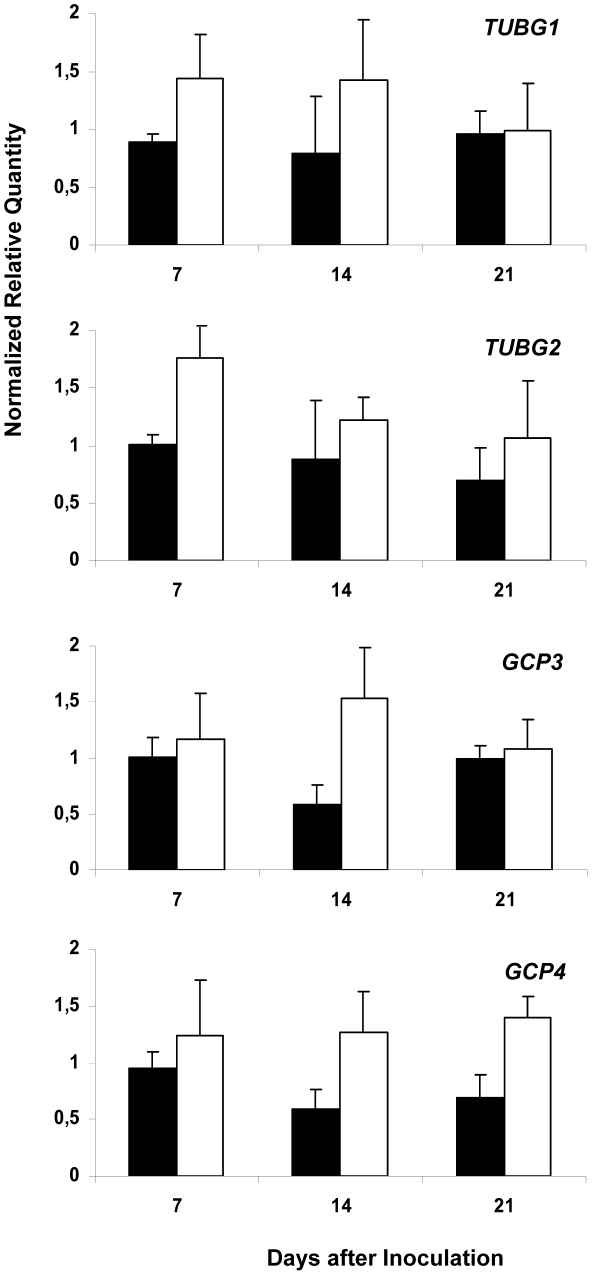
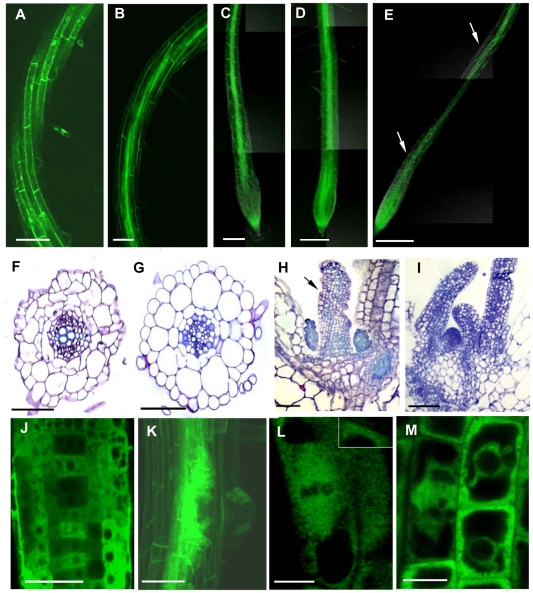
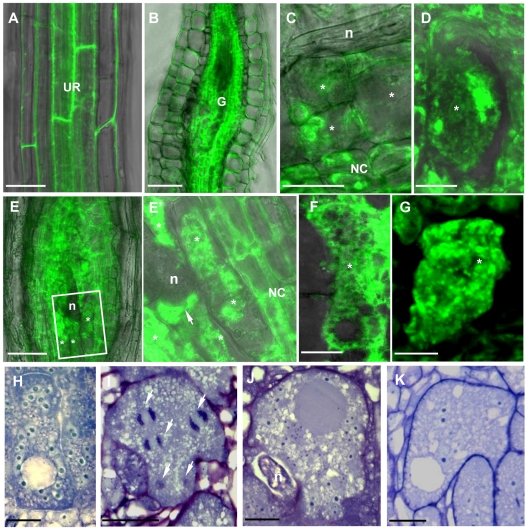
Reorganization of the microtubule network is important for the fast isodiametric expansion of giant-feeding cells induced by root-knot nematodes. The efficiency of microtubule reorganization depends on the nucleation of new microtubules, their elongation rate and activity of microtubule severing factors. New microtubules in plants are nucleated by cytoplasmic or microtubule-bound γ-tubulin ring complexes. Here we investigate the requirement of γ-tubulin complexes for giant feeding cells development using the interaction between Arabidopsis and Meloidogyne spp. as a model system. Immunocytochemical analyses demonstrate that γ-tubulin localizes to both cortical cytoplasm and mitotic microtubule arrays of the giant cells where it can associate with microtubules. The transcripts of two Arabidopsis γ-tubulin (TUBG1 and TUBG2) and two γ-tubulin complex proteins genes (GCP3 and GCP4) are upregulated in galls. Electron microscopy demonstrates association of GCP3 and γ-tubulin as part of a complex in the cytoplasm of giant cells. Knockout of either or both γ-tubulin genes results in the gene dose-dependent alteration of the morphology of feeding site and failure of nematode life cycle completion. We conclude that the γ-tubulin complex is essential for the control of microtubular network remodelling in the course of initiation and development of giant-feeding cells, and for the successful reproduction of nematodes in their plant hosts.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures






Similar articles
-
Dynamic cytoskeleton rearrangements in giant cells and syncytia of nematode-infected roots.Plant J. 2004 Apr;38(1):12-26. doi: 10.1111/j.1365-313X.2004.02019.x. Plant J. 2004. PMID: 15053756
-
Arabidopsis GCP3-interacting protein 1/MOZART 1 is an integral component of the γ-tubulin-containing microtubule nucleating complex.Plant J. 2012 Jul;71(2):216-25. doi: 10.1111/j.1365-313X.2012.04988.x. Epub 2012 May 14. Plant J. 2012. PMID: 22404201
-
Cell Wall Modifications in Giant Cells Induced by the Plant Parasitic Nematode Meloidogyne incognita in Wild-Type (Col-0) and the fra2 Arabidopsis thaliana Katanin Mutant.Int J Mol Sci. 2019 Nov 2;20(21):5465. doi: 10.3390/ijms20215465. Int J Mol Sci. 2019. PMID: 31684028 Free PMC article.
-
Manipulation of host gene expression by root-knot nematodes.J Parasitol. 1996 Dec;82(6):881-8. J Parasitol. 1996. PMID: 8973394 Review.
-
Nematode-induced endoreduplication in plant host cells: why and how?Mol Plant Microbe Interact. 2013 Jan;26(1):17-24. doi: 10.1094/MPMI-05-12-0128-CR. Mol Plant Microbe Interact. 2013. PMID: 23194340 Review.
Cited by
-
The root-knot nematode effector MiPFN3 disrupts plant actin filaments and promotes parasitism.PLoS Pathog. 2018 Mar 15;14(3):e1006947. doi: 10.1371/journal.ppat.1006947. eCollection 2018 Mar. PLoS Pathog. 2018. PMID: 29543900 Free PMC article.
-
Transcriptome profiling of the fifth-stage larvae of Angiostrongylus cantonensis by next-generation sequencing.Parasitol Res. 2013 Sep;112(9):3193-202. doi: 10.1007/s00436-013-3495-z. Epub 2013 Jul 5. Parasitol Res. 2013. PMID: 23828188 Free PMC article.
-
Discovery of two tightly linked soybean genes at the qSCN10 (O) locus conferring broad-spectrum resistance to soybean cyst nematode.Commun Biol. 2025 Feb 18;8(1):259. doi: 10.1038/s42003-025-07633-8. Commun Biol. 2025. PMID: 39966671 Free PMC article.
-
Plasmodesmata play pivotal role in sucrose supply to Meloidogyne graminicola-caused giant cells in rice.Mol Plant Pathol. 2021 May;22(5):539-550. doi: 10.1111/mpp.13042. Epub 2021 Mar 15. Mol Plant Pathol. 2021. PMID: 33723908 Free PMC article.
-
Application of Nuclear Volume Measurements to Comprehend the Cell Cycle in Root-Knot Nematode-Induced Giant Cells.Front Plant Sci. 2017 Jun 12;8:961. doi: 10.3389/fpls.2017.00961. eCollection 2017. Front Plant Sci. 2017. PMID: 28659939 Free PMC article.
References
-
- Jansky SH, Simon R, Spooner DM. A test of taxonomic predictivity: Resistance to early blight in wild relatives of cultivated potato. Phytopathology. 2008;98:680–687. - PubMed
-
- Wyss U, Grundler FMW, Munch A. The parasitic behavior of second-stage juveniles of Meloidogyne incognita in roots of Arabidopsis thaliana. Nematologica. 1992;38:98–111.
-
- Von Mende N. Invasion and migration behaviour of sedentary nematodes. In: Fenoll C, Grundler FMW, Ohl SA, editors. Cellular and molecular aspects of plant-nematodes interactions. Vol. 21. Dordrecht, The Netherlands: Kluwer Academics Publishers; 1997. pp. 1129–1140.
-
- Jones MGK. The development and function of plant cells modified by endoparasitic nematodes. In: Zuckerman BM, Rhode RA, editors. Plant Parasitic Nematodes. New York: Academic Press; 1981. pp. 225–279.
-
- Wiggers RJ, Starr JL, Price HJ. DNA content and variation in chromosome number in plant cells affected by Meloidogyne incognita and M. arenaria. Phytopathology. 1990;80:1391–1395.
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
