

Wall teichoic acids of Staphylococcus aureus limit recognition by the drosophila peptidoglycan recognition protein-SA to promote pathogenicity
- PMID: 22144903
- PMCID: PMC3228820
- DOI: 10.1371/journal.ppat.1002421
Wall teichoic acids of Staphylococcus aureus limit recognition by the drosophila peptidoglycan recognition protein-SA to promote pathogenicity
Abstract
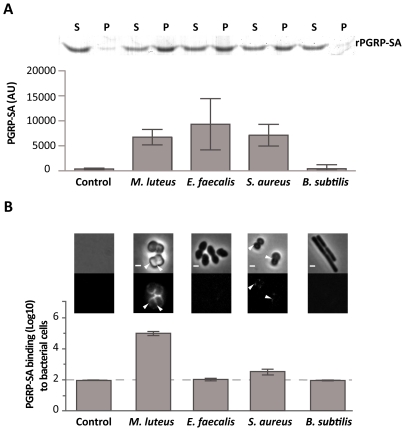
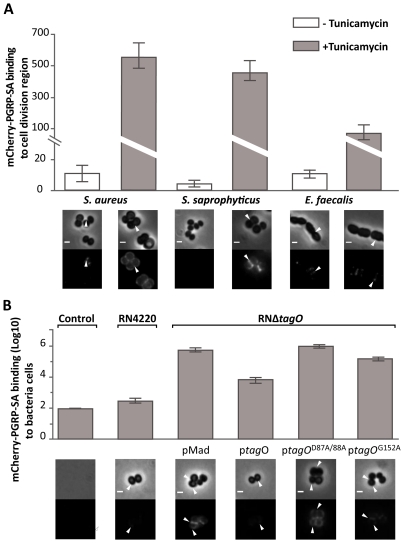
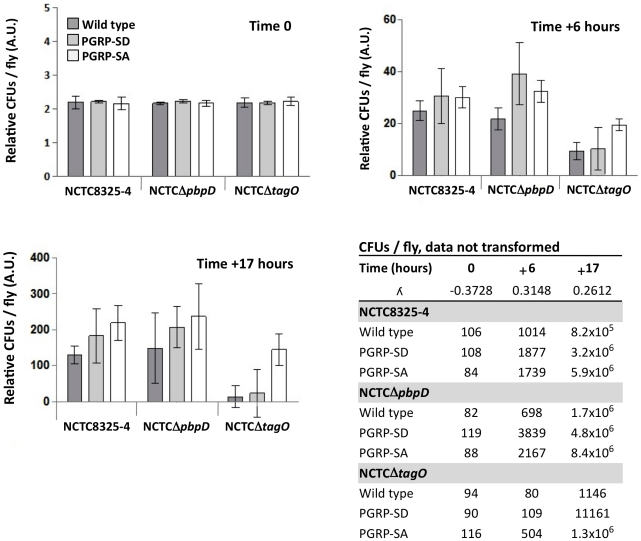
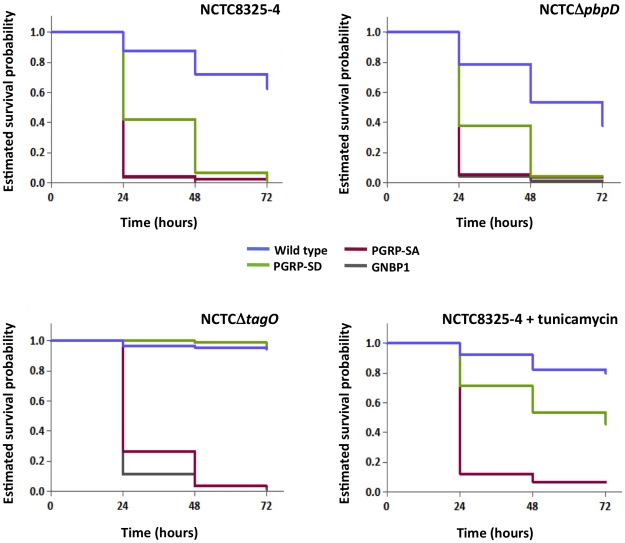
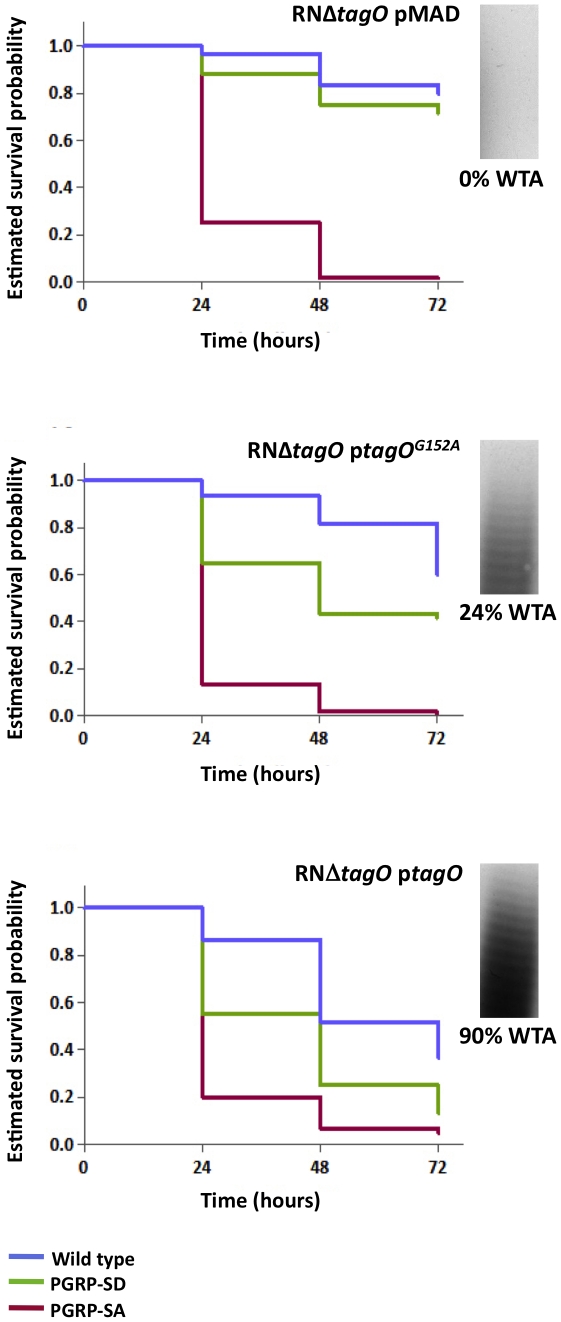
The cell wall of gram-positive bacteria is a complex network of surface proteins, capsular polysaccharides and wall teichoic acids (WTA) covalently linked to Peptidoglycan (PG). The absence of WTA has been associated with a reduced pathogenicity of Staphylococcus aureus (S. aureus). Here, we assessed whether this was due to increased detection of PG, an important target of innate immune receptors. Antibiotic-mediated or genetic inhibition of WTA production in S. aureus led to increased binding of the non-lytic PG Recognition Protein-SA (PGRP-SA), and this was associated with a reduction in host susceptibility to infection. Moreover, PGRP-SD, another innate sensor required to control wild type S. aureus infection, became redundant. Our data imply that by using WTA to limit access of innate immune receptors to PG, under-detected bacteria are able to establish an infection and ultimately overwhelm the host. We propose that different PGRPs work in concert to counter this strategy.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
Accessibility to Peptidoglycan Is Important for the Recognition of Gram-Positive Bacteria in Drosophila.Cell Rep. 2019 May 21;27(8):2480-2492.e6. doi: 10.1016/j.celrep.2019.04.103. Cell Rep. 2019. PMID: 31116990 Free PMC article.
-
Biochemical characterization of evasion from peptidoglycan recognition by Staphylococcus aureus D-alanylated wall teichoic acid in insect innate immunity.Dev Comp Immunol. 2011 Aug;35(8):835-9. doi: 10.1016/j.dci.2011.03.001. Epub 2011 Mar 29. Dev Comp Immunol. 2011. PMID: 21453720
-
Inhibitory role for D-alanylation of wall teichoic acid in activation of insect Toll pathway by peptidoglycan of Staphylococcus aureus.J Immunol. 2010 Aug 15;185(4):2424-31. doi: 10.4049/jimmunol.1000625. Epub 2010 Jul 16. J Immunol. 2010. PMID: 20639481
-
Function and regulation of Staphylococcus aureus wall teichoic acids and capsular polysaccharides.Int J Med Microbiol. 2019 Sep;309(6):151333. doi: 10.1016/j.ijmm.2019.151333. Epub 2019 Jul 18. Int J Med Microbiol. 2019. PMID: 31362856 Review.
-
The staphylococcal surface-glycopolymer wall teichoic acid (WTA) is crucial for complement activation and immunological defense against Staphylococcus aureus infection.Immunobiology. 2016 Oct;221(10):1091-101. doi: 10.1016/j.imbio.2016.06.003. Epub 2016 Jun 15. Immunobiology. 2016. PMID: 27424796 Review.
Cited by
-
MprF-mediated immune evasion is necessary for Lactiplantibacillus plantarum resilience in the Drosophila gut during inflammation.PLoS Pathog. 2024 Aug 19;20(8):e1012462. doi: 10.1371/journal.ppat.1012462. eCollection 2024 Aug. PLoS Pathog. 2024. PMID: 39159259 Free PMC article.
-
Teichoic acid biosynthesis as an antibiotic target.Curr Opin Microbiol. 2013 Oct;16(5):531-7. doi: 10.1016/j.mib.2013.06.014. Epub 2013 Jul 31. Curr Opin Microbiol. 2013. PMID: 23916223 Free PMC article. Review.
-
Structural and functional analysis of the D-alanyl carrier protein ligase DltA from Staphylococcus aureus Mu50.Acta Crystallogr D Struct Biol. 2022 Apr 1;78(Pt 4):424-434. doi: 10.1107/S2059798322000547. Epub 2022 Mar 16. Acta Crystallogr D Struct Biol. 2022. PMID: 35362466 Free PMC article.
-
Revisiting Anti-tuberculosis Therapeutic Strategies That Target the Peptidoglycan Structure and Synthesis.Front Microbiol. 2019 Feb 11;10:190. doi: 10.3389/fmicb.2019.00190. eCollection 2019. Front Microbiol. 2019. PMID: 30804921 Free PMC article. Review.
-
Accessibility to Peptidoglycan Is Important for the Recognition of Gram-Positive Bacteria in Drosophila.Cell Rep. 2019 May 21;27(8):2480-2492.e6. doi: 10.1016/j.celrep.2019.04.103. Cell Rep. 2019. PMID: 31116990 Free PMC article.
References
-
- Foster TJ. Immune evasion by staphylococci. Nat Rev Microbiol. 2005;3:948–958. - PubMed
-
- Chaput C, Boneca IG. Peptidoglycan detection by mammals and flies. Microbes Infect. 2007;9:637–647. - PubMed
-
- Vollmer W, Blanot D, de Pedro MA. Peptidoglycan structure and architecture. FEMS Microbiol Rev. 2008;32:149–167. - PubMed
-
- Scott JR, Barnett TC. Surface proteins of gram-positive bacteria and how they get there. Annu Rev Microbiol. 2006;60:397–423. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
