

LKB1 is an essential regulator of spermatozoa release during spermiation in the mammalian testis
- PMID: 22145035
- PMCID: PMC3228757
- DOI: 10.1371/journal.pone.0028306
LKB1 is an essential regulator of spermatozoa release during spermiation in the mammalian testis
Abstract
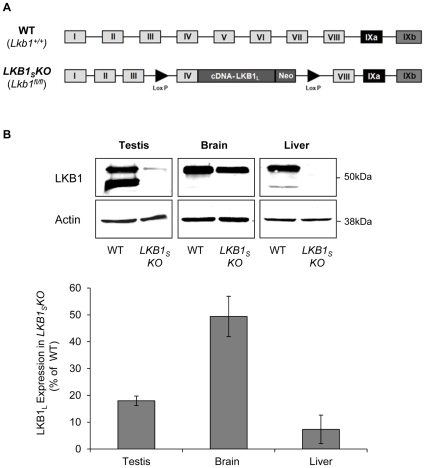
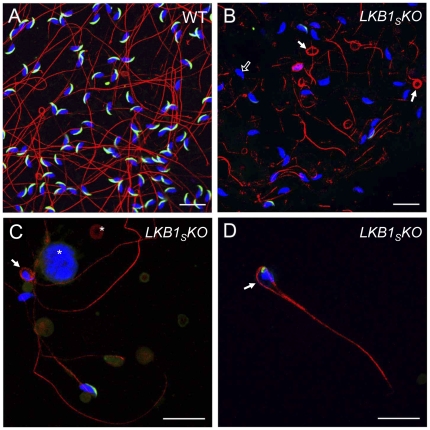
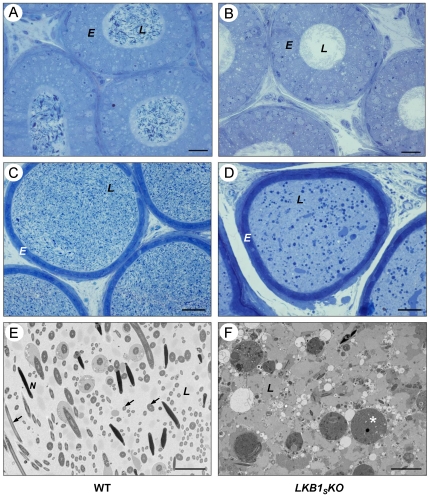
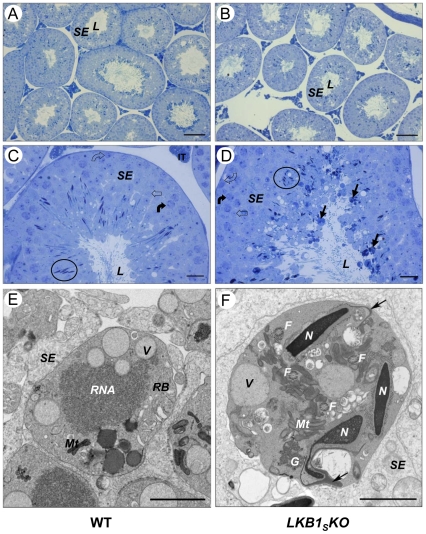
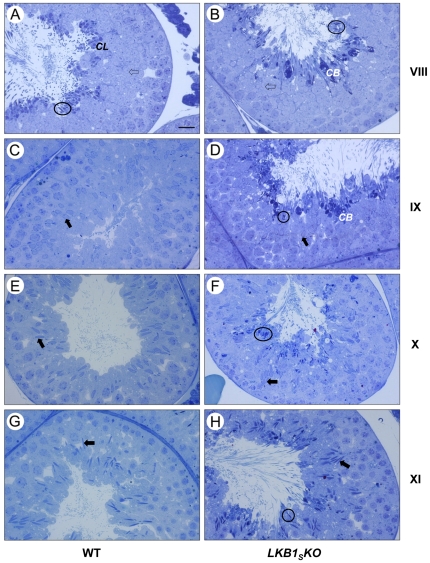
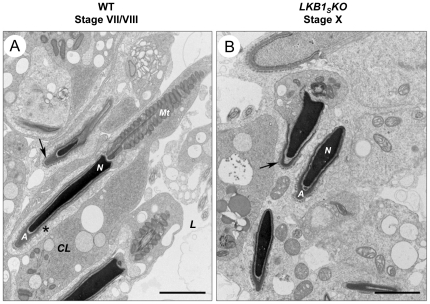
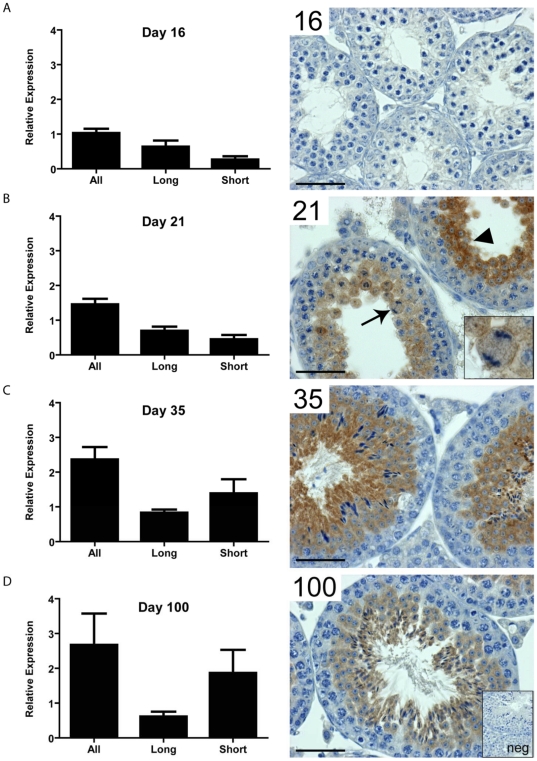
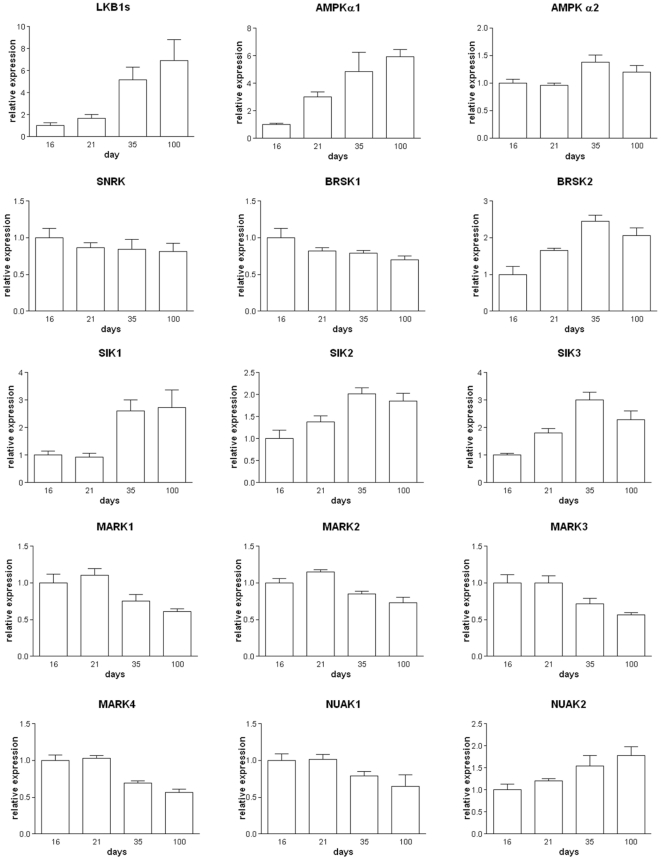
LKB1 acts as a master upstream protein kinase regulating a number of kinases involved in diverse cellular functions. Recent studies have suggested a role for LKB1 in male fertility. Male mice with reduced total LKB1 expression, including the complete absence of the major splice variant in testis (LKB1(S)), are completely infertile. We sought to further characterise these mice and determine the mechanism underlying this infertility. This involved expression studies of LKB1 in developing germ cells, morphological analysis of mature spermatozoa and histological studies of both the testis and epididymis using light microscopy and transmission electron microscopy. We conclude that a defect in the release of mature spermatids from the seminiferous epithelium (spermiation) during spermatozoan development is a major cause of the infertility phenotype. We also present evidence that this is due, at least in part, to defects in the breakdown of the junctions, known as ectoplasmic specialisations, between the sertoli cells of the testis epithelium and the heads of the maturing spermatids. Overall this study uncovers a critical role for LKB1 in spermiation, a highly regulated, but poorly understood process vital for male fertility.
Conflict of interest statement
Figures
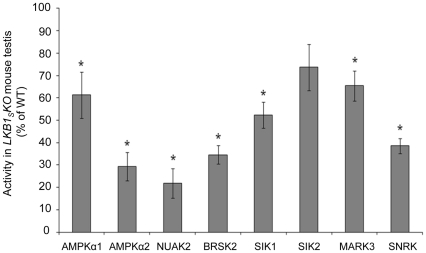
References
-
- Alessi DR, Sakamoto K, Bayascas JR. LKB1-dependent signaling pathways. Annu Rev Biochem. 2006;75:137–163. - PubMed
-
- Jenne DE, Reimann H, Nezu J, Friedel W, Loff S, et al. Peutz-Jeghers syndrome is caused by mutations in a novel serine threonine kinase. Nat Genet. 1998;18:38–43. - PubMed
-
- Hemminki A, Markie D, Tomlinson I, Avizienyte E, Roth S, et al. A serine/threonine kinase gene defective in Peutz-Jeghers syndrome. Nature. 1998;391:184–187. - PubMed
-
- Ylikorkala A, Rossi DJ, Korsisaari N, Luukko K, Alitalo K, et al. Vascular abnormalities and deregulation of VEGF in Lkb1-deficient mice. Science. 2001;293:1323–1326. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
