

Non-DNA-binding cofactors enhance DNA-binding specificity of a transcriptional regulatory complex
- PMID: 22146299
- PMCID: PMC3737730
- DOI: 10.1038/msb.2011.89
Non-DNA-binding cofactors enhance DNA-binding specificity of a transcriptional regulatory complex
Abstract
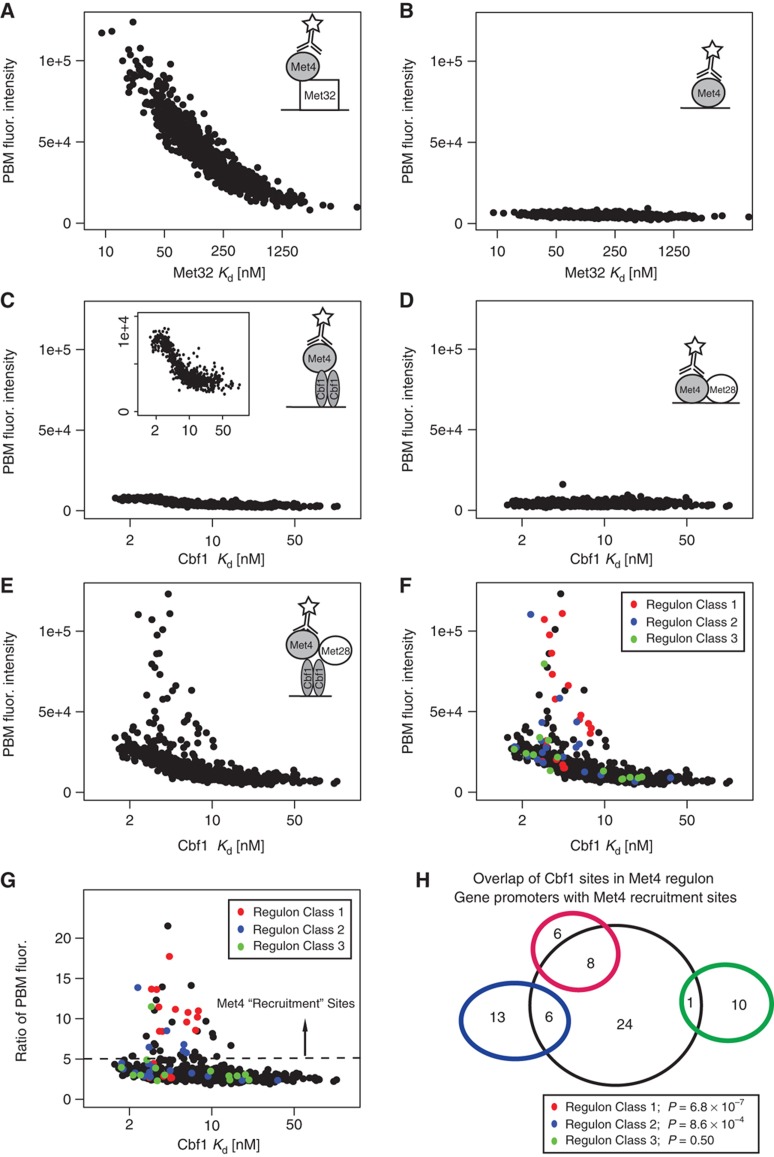
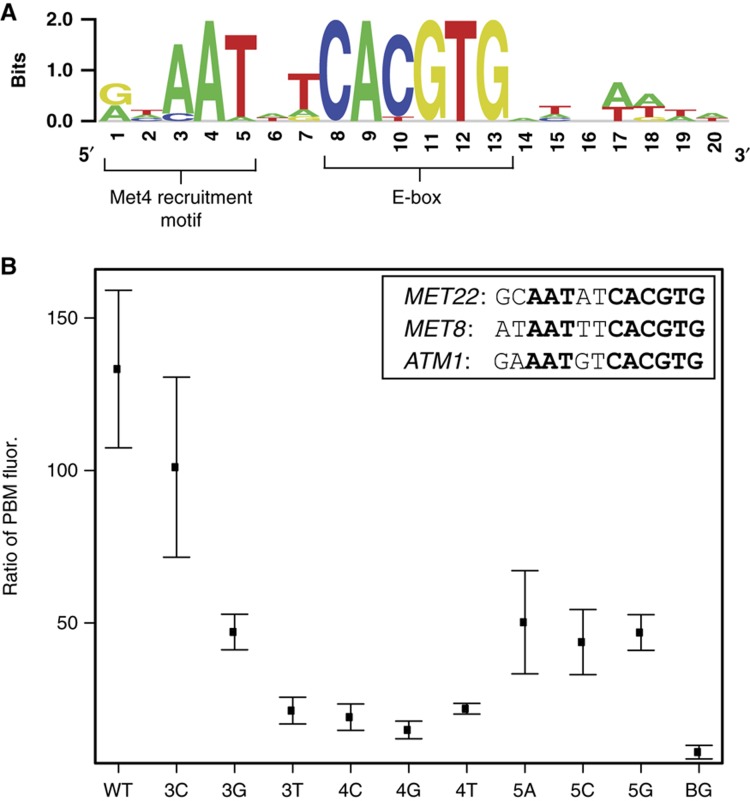
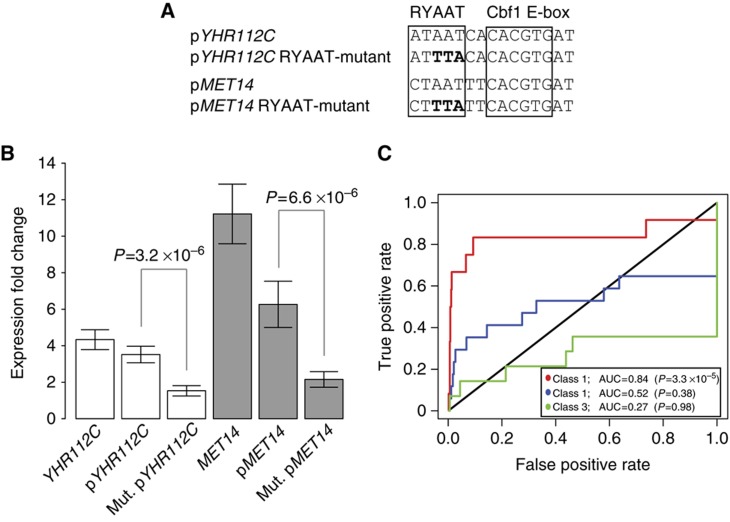
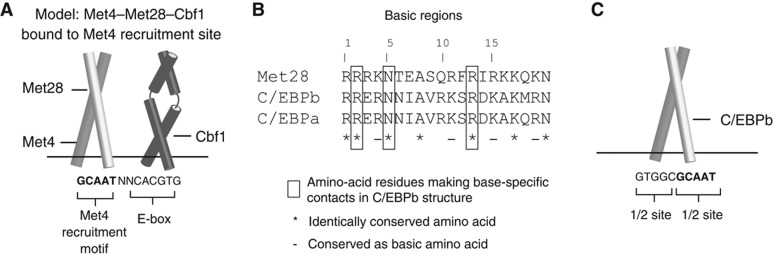
Recruitment of cofactors to specific DNA sites is integral for specificity in gene regulation. As a model system, we examined how targeting and transcriptional control of the sulfur metabolism genes in Saccharomyces cerevisiae is governed by recruitment of the transcriptional co-activator Met4. We developed genome-scale approaches to measure transcription factor (TF) DNA-binding affinities and cofactor recruitment to >1300 genomic binding site sequences. We report that genes responding to the TF Cbf1 and cofactor Met28 contain a novel 'recruitment motif' (RYAAT), adjacent to Cbf1 binding sites, which enhances the binding of a Met4-Met28-Cbf1 regulatory complex, and that abrogation of this motif significantly reduces gene induction under low-sulfur conditions. Furthermore, we show that correct recognition of this composite motif requires both non-DNA-binding cofactors Met4 and Met28. Finally, we demonstrate that the presence of an RYAAT motif next to a Cbf1 site, rather than Cbf1 binding affinity, specifies Cbf1-dependent sulfur metabolism genes. Our results highlight the need to examine TF/cofactor complexes, as novel specificity can result from cofactors that lack intrinsic DNA-binding specificity.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
Comment in
-
Gene regulation: Resolving transcription factor binding.Nat Rev Genet. 2011 Dec 29;13(2):71. doi: 10.1038/nrg3153. Nat Rev Genet. 2011. PMID: 22207166 No abstract available.
References
-
- Badis G, Chan ET, van Bakel H, Pena-Castillo L, Tillo D, Tsui K, Carlson CD, Gossett AJ, Hasinoff MJ, Warren CL, Gebbia M, Talukder S, Yang A, Mnaimneh S, Terterov D, Coburn D, Li Yeo A, Yeo ZX, Clarke ND, Lieb JD et al. (2008) A library of yeast transcription factor motifs reveals a widespread function for Rsc3 in targeting nucleosome exclusion at promoters. Mol Cell 32: 878–887 - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
