

Review
doi: 10.1104/pp.111.189845.
Epub 2011 Dec 6.
Natural variation in Arabidopsis: from molecular genetics to ecological genomics
Affiliations
- PMID: 22147517
- PMCID: PMC3252104
- DOI: 10.1104/pp.111.189845
Item in Clipboard
Review
Natural variation in Arabidopsis: from molecular genetics to ecological genomics
Plant Physiol.
2012 Jan.
No abstract available
Figures

Gross morphological variation in Arabidopsis and relatives. A, Variation between Arabidopsis accessions. On top, vegetative rosettes of accessions grown for 4 weeks in long days are shown. They vary in rosette diameter and compactness, leaf shape, and tissue necrosis or onset of senescence. Similarly, variation in size and shape of individual leaves, in this case the sixth in the rosette, is apparent in the 10 examples shown on the bottom left. Finally, differences in overall architecture are illustrated with five plants. On the left is an early flowering accession with few rosette leaves. The next two flower later, but the second one from the left has reduced apical dominance. Finally, the two accessions on the right have similarly tall main inflorescences but differ in the number of secondary inflorescences. The appearance on the far right is common among wild-grown plants. B, Some characters, such as flower size and fruit shape, vary relatively little within Arabidopsis, but more dramatic variation is found in comparison with closely related taxa, such as Capsella rubella (left) and A. lyrata (right). Images courtesy of Eunyoung Chae, Sang-Tae Kim, and George Wang.

Populations for mapping genes causing trait variation. Colors indicate contribution from different parental accessions. Only one chromosome pair is shown for each individual. HIF individuals are derived from RILs, in which a small portion of the genome is still heterozygous.
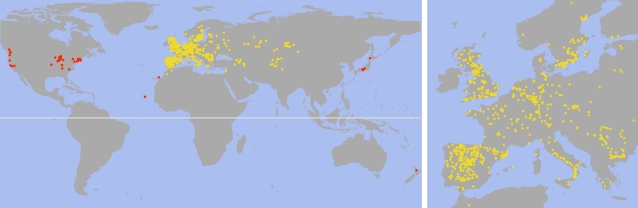
Distribution of over 7,000 Arabidopsis accessions collected from the wild and available in the stock center or soon-to-be-released collections. Western and southern Europe, including Great Britain, is heavily overrepresented, although sampling is not even. Accessions from the presumed native range are in yellow and likely introductions in red. Whether the distribution across China to Japan is continuous with the native range is unclear. Arabidopsis has been reported in additional locales, such as South Korea, and several African countries (Alonso-Blanco and Koornneef, 2000). Maps courtesy of George Wang.
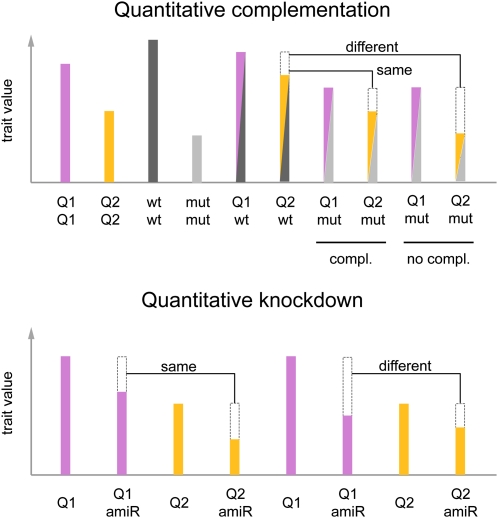
Quantitative complementation and knockdown to determine whether QTL are allelic to a candidate gene. Both tests rely on quantitative comparisons between genotypes; the dashed boxes indicate phenotypic differences to the genotype to the left. In a quantitative complementation test, one determines whether the two QTL alleles, Q1 and Q2, are differentially affected when heterozygous with the wild-type (wt) or mutant (mut) allele of a candidate gene (Mackay, 2001). If the QTL alleles respond differently, i.e. if in this example only Q1 complements the mutant phenotype, the candidate gene and the QTL are probably allelic. Similarly, in a quantitative knockdown experiment, a differential effect of an amiRNA (amiR) against the candidate gene indicates that the Q1 allele has lower activity than Q2 and that the candidate gene is likely responsible for the QTL.
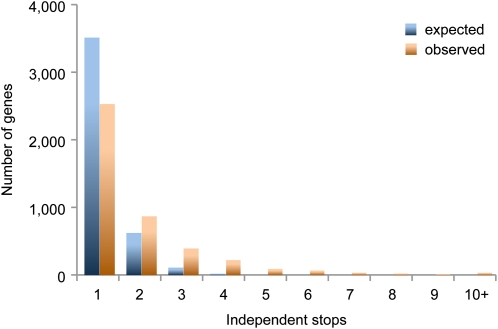
Comparison of expected and observed occurrences of 8,133 independent premature stops in 4,263 protein coding genes, considering all genes with >90% coverage in 75 out of 80 accessions. Data are from Cao et al. (2011).
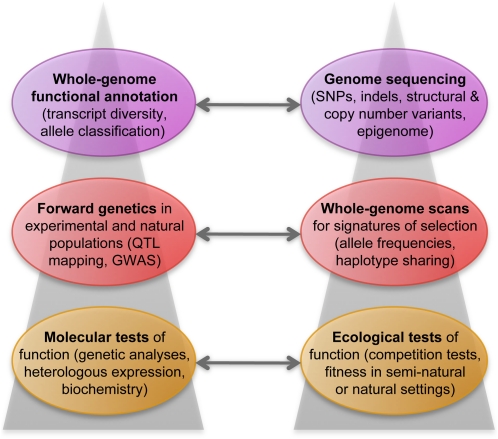
Relationship between approaches to the study of genetic variation.
Comment in
-
Founders review 2012.Plant Physiol. 2012 Jan;158(1):1. doi: 10.1104/pp.111.900428. Plant Physiol. 2012. PMID: 22213247 Free PMC article. No abstract available.
References
-
- Adams S, Allen T, Whitelam GC. (2009) Interaction between the light quality and flowering time pathways in Arabidopsis. Plant J 60: 257–267 - PubMed
-
- Alcázar R, García AV, Kronholm I, de Meaux J, Koornneef M, Parker JE, Reymond M. (2010) Natural variation at Strubbelig Receptor Kinase 3 drives immune-triggered incompatibilities between Arabidopsis thaliana accessions. Nat Genet 42: 1135–1139 - PubMed
-
- Alonso JM, Ecker JR. (2006) Moving forward in reverse: genetic technologies to enable genome-wide phenomic screens in Arabidopsis. Nat Rev Genet 7: 524–536 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
