

The muscle pattern of the Drosophila abdomen depends on a subdivision of the anterior compartment of each segment
- PMID: 22147953
- PMCID: PMC3231773
- DOI: 10.1242/dev.073692
The muscle pattern of the Drosophila abdomen depends on a subdivision of the anterior compartment of each segment
Abstract
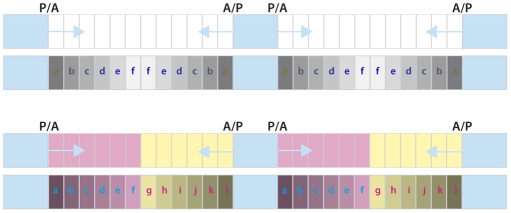
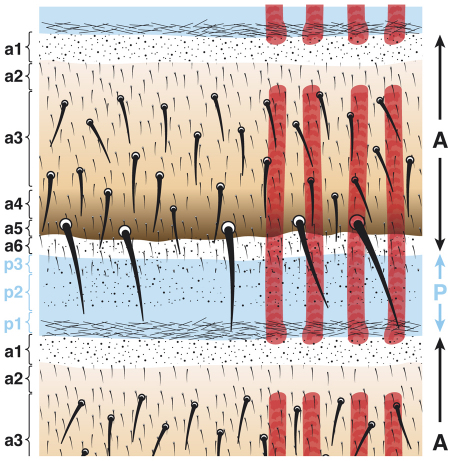
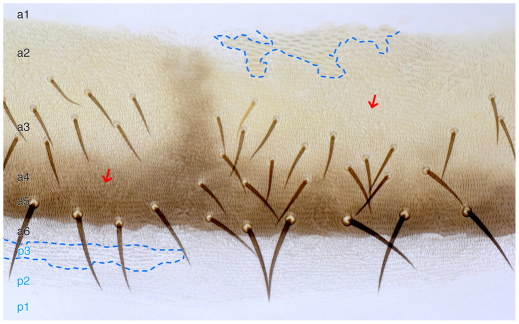
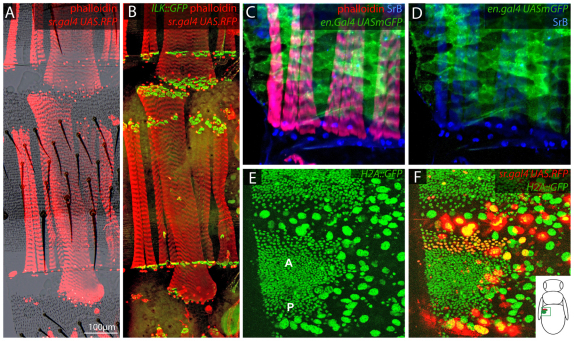
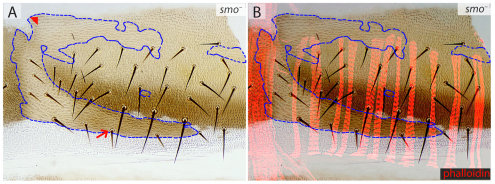
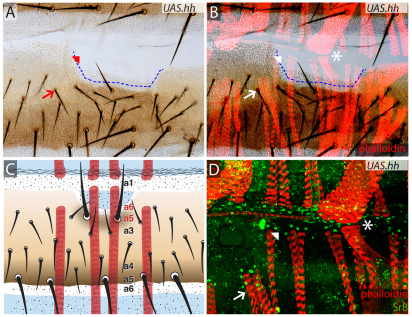
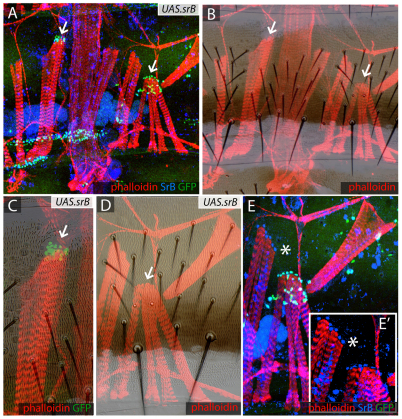
In the past, segments were defined by landmarks such as muscle attachments, notably by Snodgrass, the king of insect anatomists. Here, we show how an objective definition of a segment, based on developmental compartments, can help explain the dorsal abdomen of adult Drosophila. The anterior (A) compartment of each segment is subdivided into two domains of cells, each responding differently to Hedgehog. The anterior of these domains is non-neurogenic and clones lacking Notch develop normally; this domain can express stripe and form muscle attachments. The posterior domain is neurogenic and clones lacking Notch do not form cuticle; this domain is unable to express stripe or form muscle attachments. The posterior (P) compartment does not form muscle attachments. Our in vivo films indicate that early in the pupa the anterior domain of the A compartment expresses stripe in a narrowing zone that attracts the extending myotubes and resolves into the attachment sites for the dorsal abdominal muscles. We map the tendon cells precisely and show that all are confined to the anterior domain of A. It follows that the dorsal abdominal muscles are intersegmental, spanning from one anterior domain to the next. This view is tested and supported by clones that change cell identity or express stripe ectopically. It seems that growing myotubes originate in posterior A and extend forwards and backwards until they encounter and attach to anterior A cells. The dorsal adult muscles are polarised in the anteroposterior axis: we disprove the hypothesis that muscle orientation depends on genes that define planar cell polarity in the epidermis.
Figures

References
-
- Akam M. (1987). The molecular basis for metameric pattern in the Drosophila embryo. Development 101, 1–22 - PubMed
-
- Bate M. (1990). The embryonic development of larval muscles in Drosophila. Development 110, 791–804 - PubMed
-
- Bate M., Rushton E., Currie D. A. (1991). Cells with persistent twist expression are the embryonic precursors of adult muscles in Drosophila. Development 113, 79–89 - PubMed
-
- Becker S., Pasca G., Strumpf D., Min L., Volk T. (1997). Reciprocal signaling between Drosophila epidermal muscle attachment cells and their corresponding muscles. Development 124, 2615–2622 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
