

Zinc supplementation alters airway inflammation and airway hyperresponsiveness to a common allergen
- PMID: 22151973
- PMCID: PMC3250936
- DOI: 10.1186/1476-9255-8-36
Zinc supplementation alters airway inflammation and airway hyperresponsiveness to a common allergen
Abstract
Background: Zinc supplementation can modulate immunity through inhibition of NF-κB, a transcription factor that controls many immune response genes. Thus, we sought to examine the mechanism by which zinc supplementation tempers the response to a common allergen and determine its effect on allergic airway inflammation.
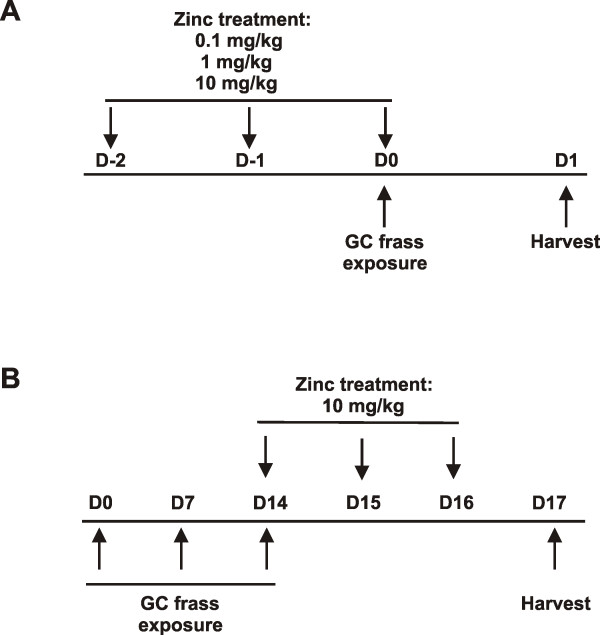
Methods: Mice were injected with zinc gluconate prior to German cockroach (GC) feces (frass) exposure and airway inflammation was assessed. Primary bone marrow-derived neutrophils and DMSO-differentiated HL-60 cells were used to assess the role of zinc gluconate on tumor necrosis factor (TNF)α expression. NF-κB:DNA binding and IKK activity were assessed by EMSA and in vitro kinase assay. Protein levels of A20, RIP1 and TRAF6 were assessed by Western blot analysis. Establishment of allergic airway inflammation with GC frass was followed by administration of zinc gluconate. Airway hyperresponsiveness, serum IgE levels, eosinophilia and Th2 cytokine production were assessed.
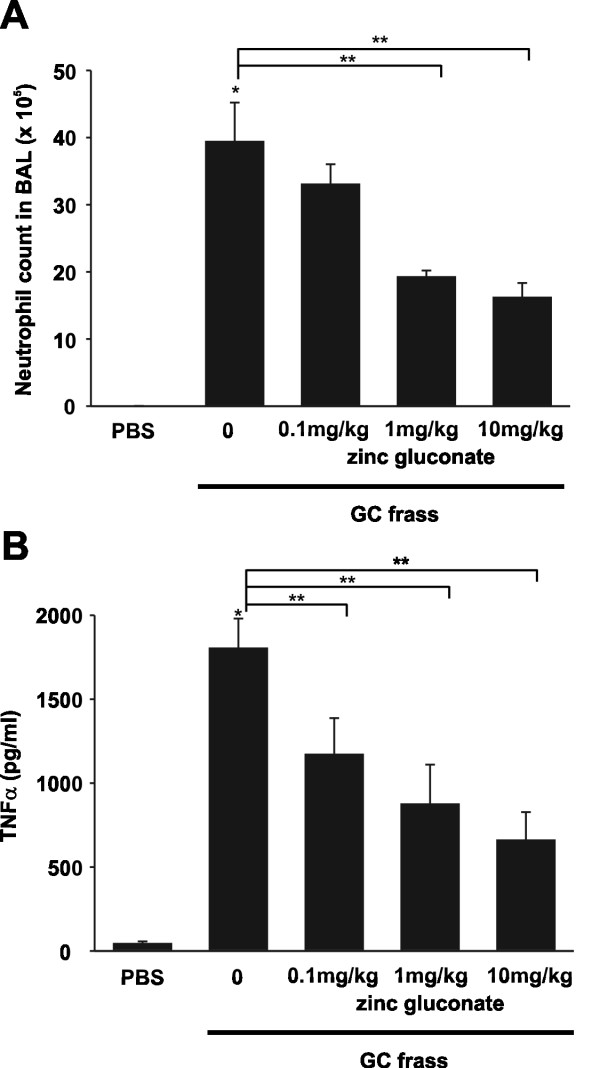
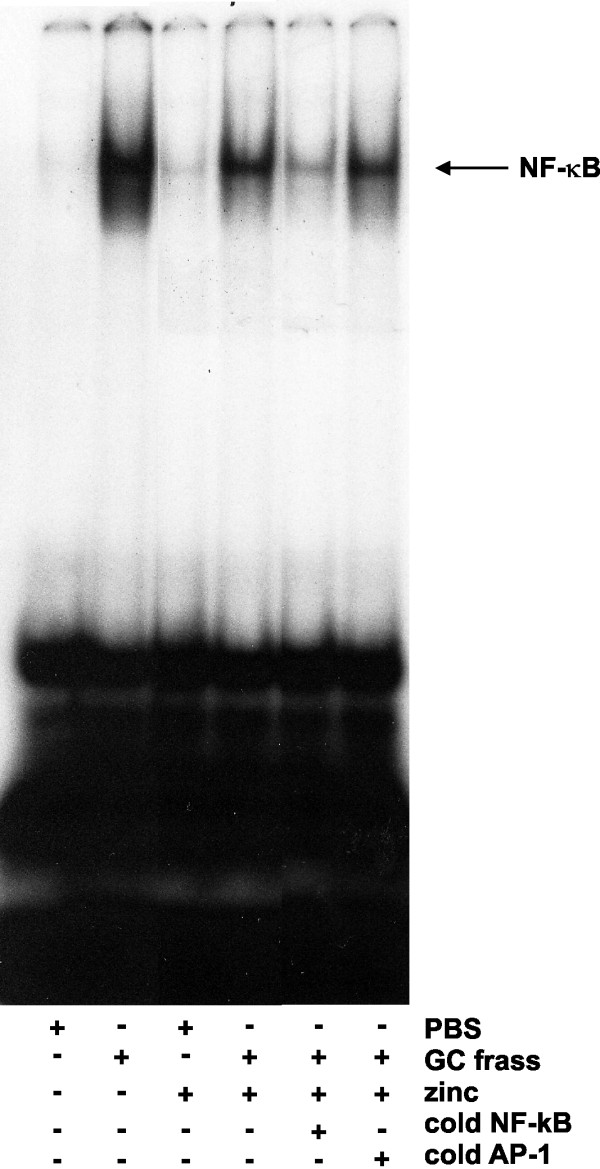
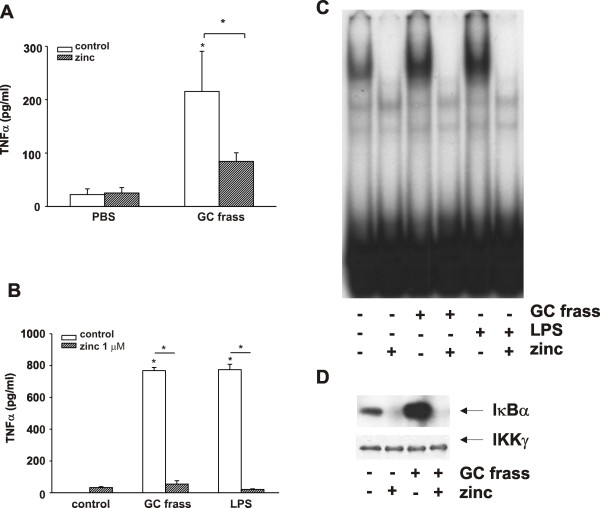
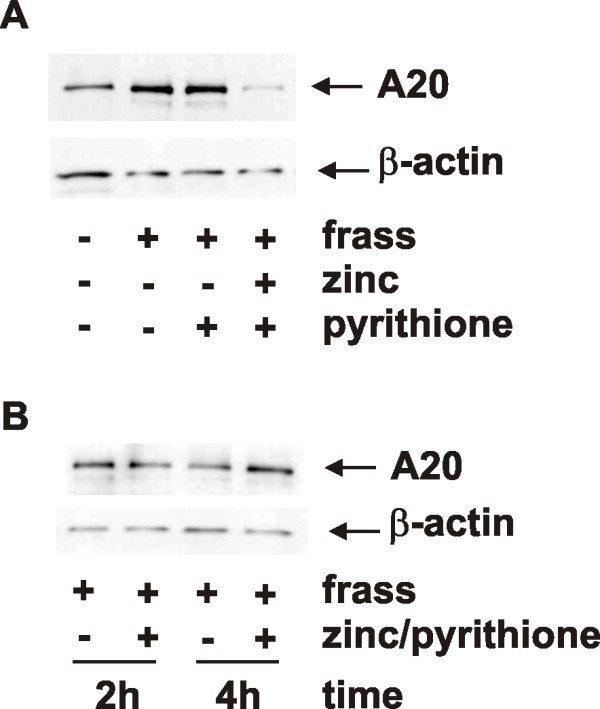
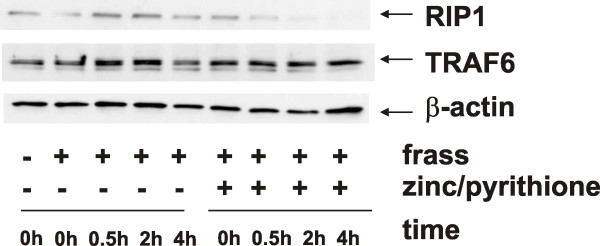
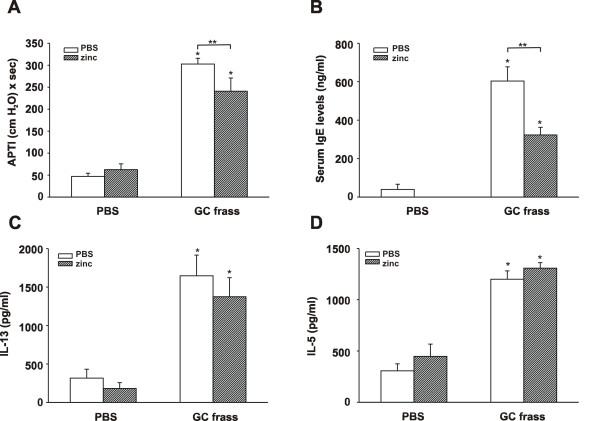
Results: Administration of zinc gluconate prior to allergen exposure resulted in significantly decreased neutrophil infiltration and TNFα cytokine release into the airways. This correlated with decreased NF-κB activity in the whole lung. Treatment with zinc gluconate significantly decreased GC frass-mediated TNFα production from bone-marrow derived neutrophils and HL-60 cells. We confirmed zinc-mediated decreases in NF-κB:DNA binding and IKK activity in HL-60 cells. A20, a natural inhibitor of NF-κB and a zinc-fingered protein, is a potential target of zinc. Zinc treatment did not alter A20 levels in the short term, but resulted in the degradation of RIP1, an important upstream activator of IKK. TRAF6 protein levels were unaffected. To determine the application for zinc as a therapeutic for asthma, we administered zinc following the establishment of allergic airway inflammation in a murine model. Zinc supplementation decreased airway hyperresponsiveness and serum IgE levels, but had no effect on Th2 cytokine expression.
Conclusions: This report suggests that the mechanism by which zinc supplementation alters NF-κB activity is via the alteration of A20 activity. In addition, this study provides evidence that supplementation of zinc to asthmatics may alter airway reactivity and serum IgE levels, suggesting zinc supplementation as a potential treatment for asthmatics.
Figures
Similar articles
-
Mucosal sensitization to German cockroach involves protease-activated receptor-2.Respir Res. 2010 May 24;11(1):62. doi: 10.1186/1465-9921-11-62. Respir Res. 2010. PMID: 20497568 Free PMC article.
-
Protease-activated receptor 2 activation of myeloid dendritic cells regulates allergic airway inflammation.Respir Res. 2011 Sep 21;12(1):122. doi: 10.1186/1465-9921-12-122. Respir Res. 2011. PMID: 21936897 Free PMC article.
-
A TLR2 agonist in German cockroach frass activates MMP-9 release and is protective against allergic inflammation in mice.J Immunol. 2009 Sep 1;183(5):3400-8. doi: 10.4049/jimmunol.0900838. Epub 2009 Aug 10. J Immunol. 2009. PMID: 19667087 Free PMC article.
-
Role of IgE in the development of allergic airway inflammation and airway hyperresponsiveness--a murine model.Allergy. 1999 Apr;54(4):297-305. doi: 10.1034/j.1398-9995.1999.00085.x. Allergy. 1999. PMID: 10371087 Review.
-
Zinc in human health: effect of zinc on immune cells.Mol Med. 2008 May-Jun;14(5-6):353-7. doi: 10.2119/2008-00033.Prasad. Mol Med. 2008. PMID: 18385818 Free PMC article. Review.
Cited by
-
Zinc, aging, and immunosenescence: an overview.Pathobiol Aging Age Relat Dis. 2015 Feb 5;5:25592. doi: 10.3402/pba.v5.25592. eCollection 2015. Pathobiol Aging Age Relat Dis. 2015. PMID: 25661703 Free PMC article.
-
Serum Zinc Status and Its Association with Allergic Sensitization: The Fifth Korea National Health and Nutrition Examination Survey.Sci Rep. 2017 Oct 3;7(1):12637. doi: 10.1038/s41598-017-13068-x. Sci Rep. 2017. PMID: 28974756 Free PMC article.
-
Antioxidant and Immune-Related Implications of Minerals in COVID-19: A Possibility for Disease Prevention and Management.Antioxidants (Basel). 2023 May 16;12(5):1104. doi: 10.3390/antiox12051104. Antioxidants (Basel). 2023. PMID: 37237970 Free PMC article. Review.
-
Immunopharmacological perspective on zinc in SARS-CoV-2 infection.Int Immunopharmacol. 2021 Jul;96:107630. doi: 10.1016/j.intimp.2021.107630. Epub 2021 Apr 1. Int Immunopharmacol. 2021. PMID: 33882442 Free PMC article. Review.
-
Potential molecular mechanisms of zinc- and copper-mediated antiviral activity on COVID-19.Nutr Res. 2021 Aug;92:109-128. doi: 10.1016/j.nutres.2021.05.008. Epub 2021 Jun 13. Nutr Res. 2021. PMID: 34284268 Free PMC article. Review.
References
-
- Shankar AH, Prasad AS. Zinc and immune function: the biological basis of altered resistance to infection. Am J Clin Nutr. 1998;68:447–463. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
