

Rotavirus nonstructural protein 1 antagonizes innate immune response by interacting with retinoic acid inducible gene I
- PMID: 22152002
- PMCID: PMC3254192
- DOI: 10.1186/1743-422X-8-526
Rotavirus nonstructural protein 1 antagonizes innate immune response by interacting with retinoic acid inducible gene I
Abstract
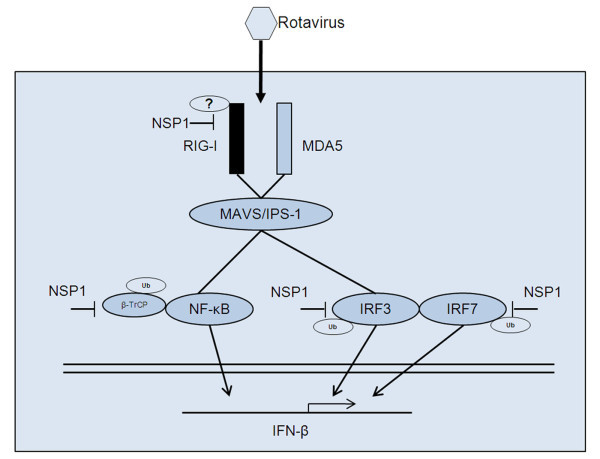
Background: The nonstructural protein 1 (NSP1) of rotavirus has been reported to block interferon (IFN) signaling by mediating proteasome-dependent degradation of IFN-regulatory factors (IRFs) and (or) the β-transducin repeat containing protein (β-TrCP). However, in addition to these targets, NSP1 may subvert innate immune responses via other mechanisms.
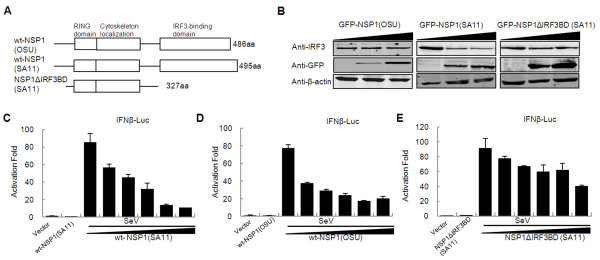
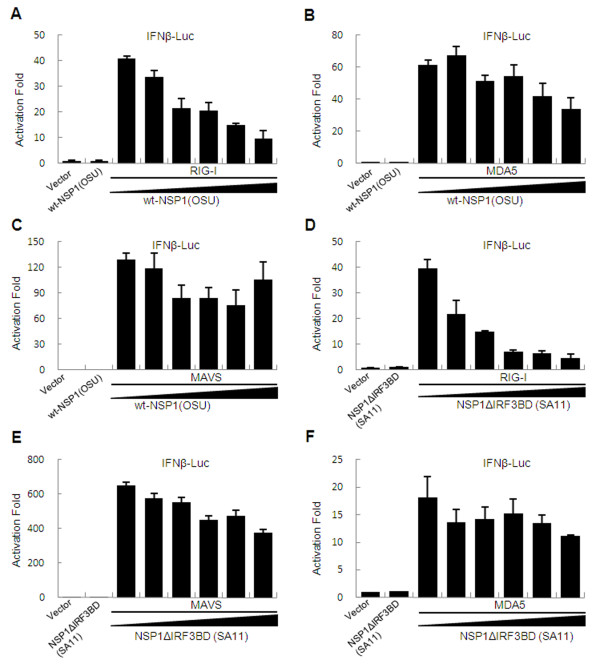
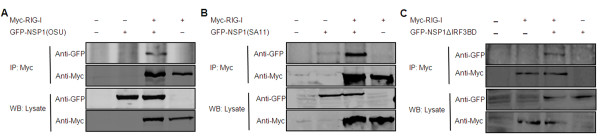
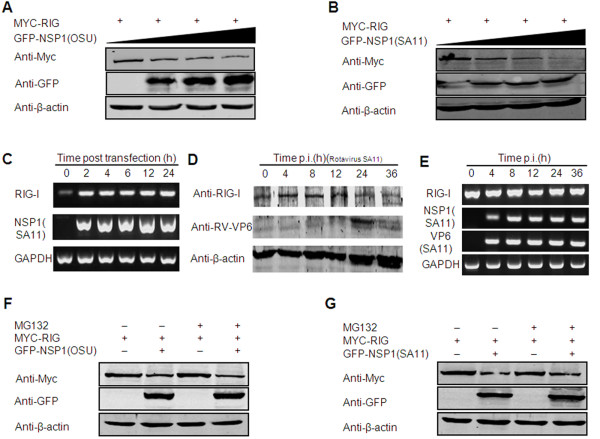
Results: The NSP1 of rotavirus OSU strain as well as the IRF3 binding domain truncated NSP1 of rotavirus SA11 strain are unable to degrade IRFs, but can still inhibit host IFN response, indicating that NSP1 may target alternative host factor(s) other than IRFs. Overexpression of NSP1 can block IFN-β promoter activation induced by the retinoic acid inducible gene I (RIG-I), but does not inhibit IFN-β activation induced by the mitochondrial antiviral-signaling protein (MAVS), indicating that NSP1 may target RIG-I. Immunoprecipitation experiments show that NSP1 interacts with RIG-I independent of IRF3 binding domain. In addition, NSP1 induces down-regulation of RIG-I in a proteasome-independent way.
Conclusions: Our findings demonstrate that inhibition of RIG-I mediated type I IFN responses by NSP1 may contribute to the immune evasion of rotavirus.
Figures
Similar articles
-
Rotavirus NSP1 inhibits NFkappaB activation by inducing proteasome-dependent degradation of beta-TrCP: a novel mechanism of IFN antagonism.PLoS Pathog. 2009 Jan;5(1):e1000280. doi: 10.1371/journal.ppat.1000280. Epub 2009 Jan 30. PLoS Pathog. 2009. PMID: 19180189 Free PMC article.
-
Diversity of interferon antagonist activities mediated by NSP1 proteins of different rotavirus strains.J Virol. 2011 Mar;85(5):1970-9. doi: 10.1128/JVI.01801-10. Epub 2010 Dec 22. J Virol. 2011. PMID: 21177809 Free PMC article.
-
Rotavirus nonstructural protein 1 subverts innate immune response by inducing degradation of IFN regulatory factor 3.Proc Natl Acad Sci U S A. 2005 Mar 15;102(11):4114-9. doi: 10.1073/pnas.0408376102. Epub 2005 Mar 1. Proc Natl Acad Sci U S A. 2005. PMID: 15741273 Free PMC article.
-
Rotavirus and reovirus modulation of the interferon response.J Interferon Cytokine Res. 2009 Sep;29(9):559-67. doi: 10.1089/jir.2009.0072. J Interferon Cytokine Res. 2009. PMID: 19694545 Free PMC article. Review.
-
Silencing the alarms: Innate immune antagonism by rotavirus NSP1 and VP3.Virology. 2015 May;479-480:75-84. doi: 10.1016/j.virol.2015.01.006. Epub 2015 Feb 25. Virology. 2015. PMID: 25724417 Free PMC article. Review.
Cited by
-
Characterization of rotavirus RNAs that activate innate immune signaling through the RIG-I-like receptors.PLoS One. 2013 Jul 23;8(7):e69825. doi: 10.1371/journal.pone.0069825. Print 2013. PLoS One. 2013. PMID: 23894547 Free PMC article.
-
Rotavirus NSP1 Subverts the Antiviral Oligoadenylate Synthetase-RNase L Pathway by Inducing RNase L Degradation.mBio. 2022 Dec 20;13(6):e0299522. doi: 10.1128/mbio.02995-22. Epub 2022 Nov 22. mBio. 2022. PMID: 36413023 Free PMC article.
-
The Role of Nucleic Acid Sensing in Controlling Microbial and Autoimmune Disorders.Int Rev Cell Mol Biol. 2019;345:35-136. doi: 10.1016/bs.ircmb.2018.08.002. Epub 2018 Sep 25. Int Rev Cell Mol Biol. 2019. PMID: 30904196 Free PMC article. Review.
-
The battle between rotavirus and its host for control of the interferon signaling pathway.PLoS Pathog. 2013 Jan;9(1):e1003064. doi: 10.1371/journal.ppat.1003064. Epub 2013 Jan 24. PLoS Pathog. 2013. PMID: 23359266 Free PMC article. Review.
-
Comparative Structure and Function Analysis of the RIG-I-Like Receptors: RIG-I and MDA5.Front Immunol. 2019 Jul 17;10:1586. doi: 10.3389/fimmu.2019.01586. eCollection 2019. Front Immunol. 2019. PMID: 31379819 Free PMC article. Review.
References
-
- Estes MKK. In: A. Z. in fields virology. Knipe DM, et al, editor. Lippincott Williams & Wilkins/Wolters Kluwer, Philadelphia; 2006. pp. 1917–1974.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
