

TGR5 activation inhibits atherosclerosis by reducing macrophage inflammation and lipid loading
- PMID: 22152303
- PMCID: PMC3627293
- DOI: 10.1016/j.cmet.2011.11.006
TGR5 activation inhibits atherosclerosis by reducing macrophage inflammation and lipid loading
Abstract
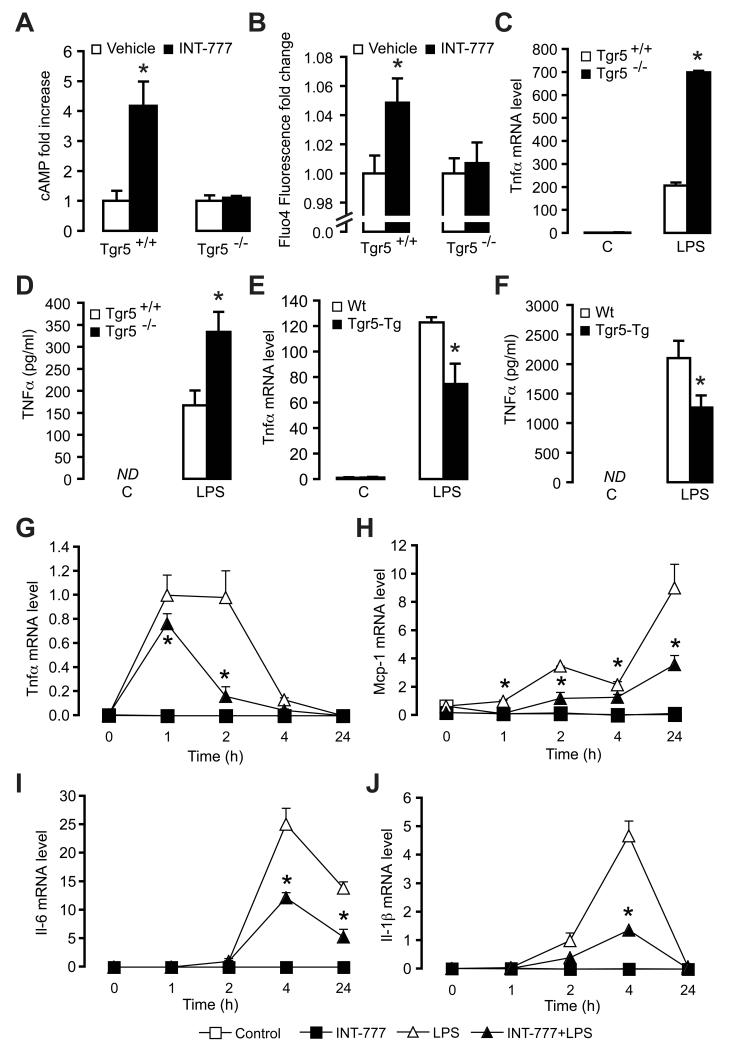
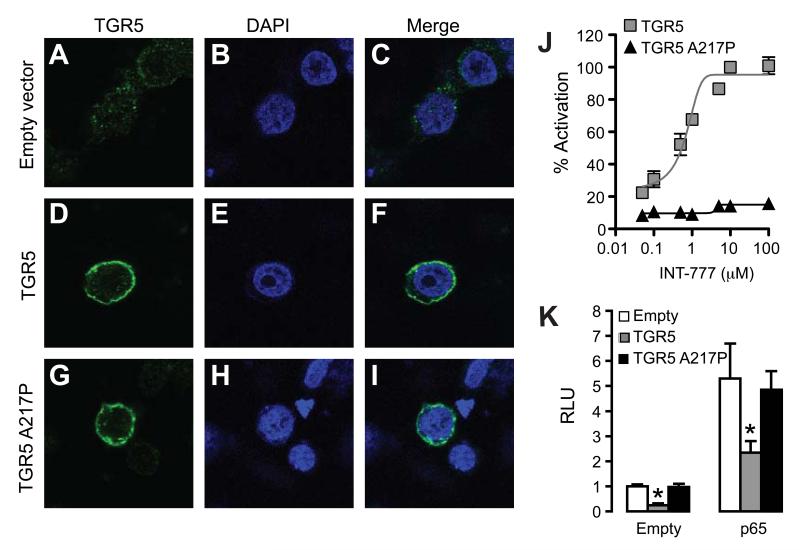
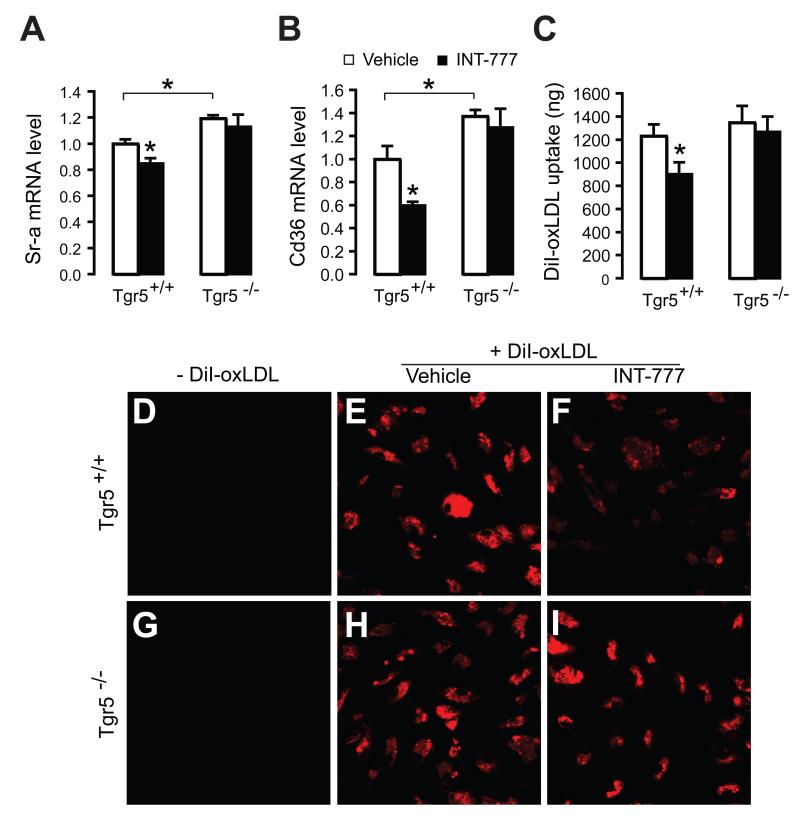
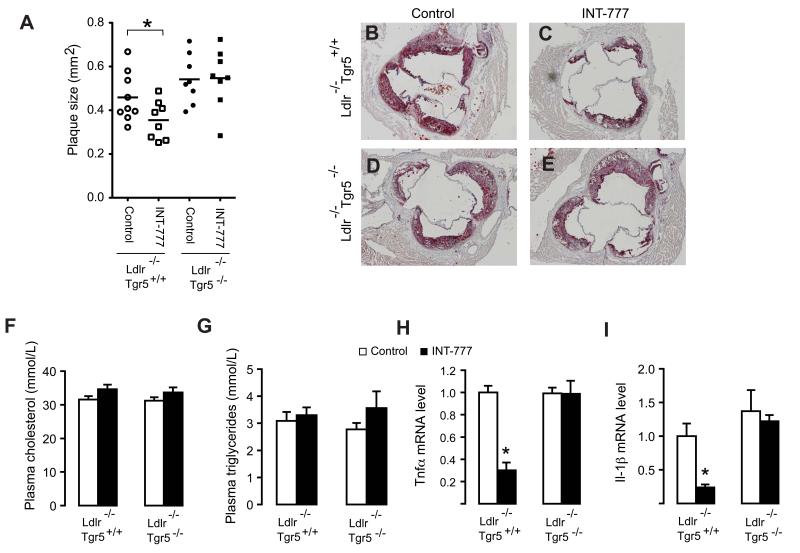
The G protein-coupled receptor TGR5 has been identified as an important component of the bile acid signaling network, and its activation has been linked to enhanced energy expenditure and improved glycemic control. Here, we demonstrate that activation of TGR5 in macrophages by 6α-ethyl-23(S)-methylcholic acid (6-EMCA, INT-777), a semisynthetic BA, inhibits proinflammatory cytokine production, an effect mediated by TGR5-induced cAMP signaling and subsequent NF-κB inhibition. TGR5 activation attenuated atherosclerosis in Ldlr(-/-)Tgr5(+/+) mice but not in Ldlr(-/-)Tgr5(-/-) double-knockout mice. The inhibition of lesion formation was associated with decreased intraplaque inflammation and less plaque macrophage content. Furthermore, Ldlr(-/-) animals transplanted with Tgr5(-/-) bone marrow did not show an inhibition of atherosclerosis by INT-777, further establishing an important role of leukocytes in INT-777-mediated inhibition of vascular lesion formation. Taken together, these data attribute a significant immune modulating function to TGR5 activation in the prevention of atherosclerosis, an important facet of the metabolic syndrome.
Copyright © 2011 Elsevier Inc. All rights reserved.
Figures

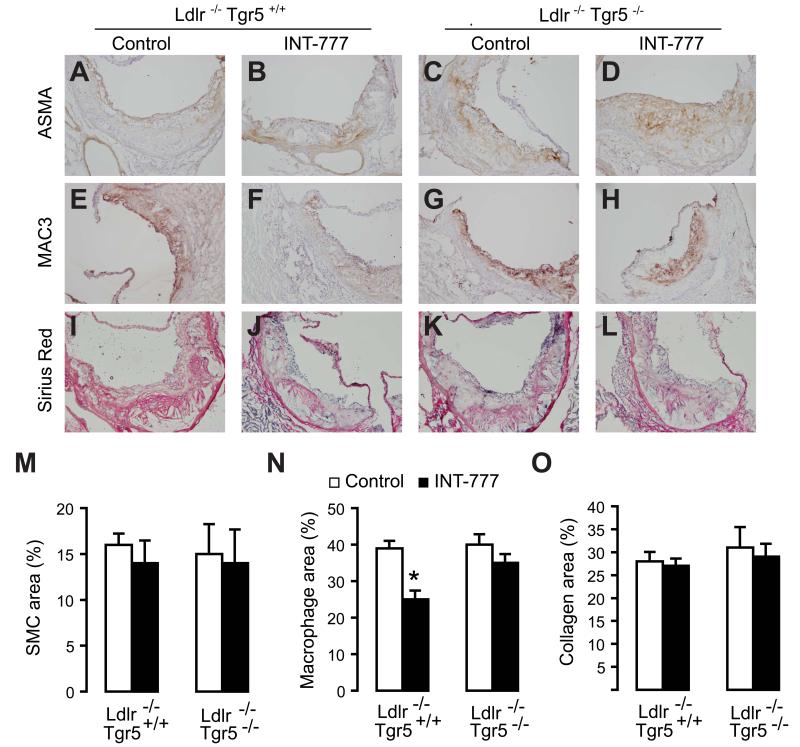
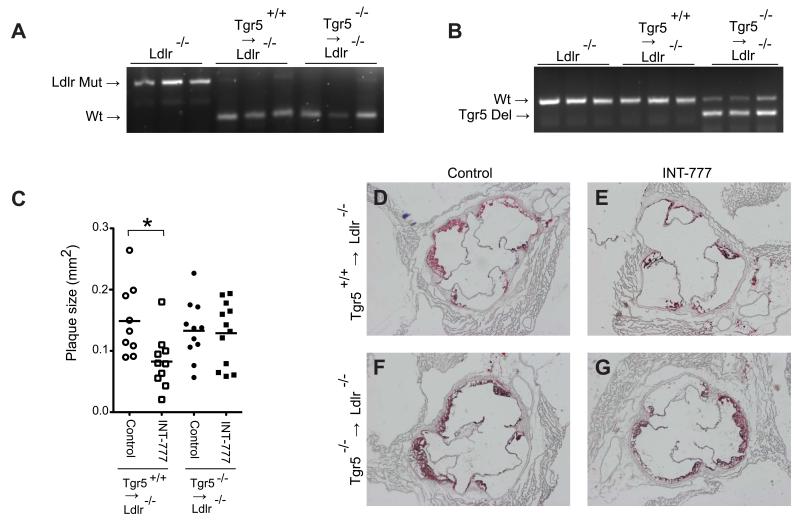
Comment in
-
G protein-coupled receptors: Case builds for TGR5 as metabolic syndrome target.Nat Rev Drug Discov. 2012 Jan 3;11(1):22. doi: 10.1038/nrd3638. Nat Rev Drug Discov. 2012. PMID: 22212676 No abstract available.
References
-
- Evans KA, Budzik BW, Ross SA, Wisnoski DD, Jin J, Rivero RA, Vimal M, Szewczyk GR, Jayawickreme C, Moncol DL, et al. Discovery of 3-aryl-4- isoxazolecarboxamides as TGR5 receptor agonists. J Med Chem. 2009;52:7962–7965. - PubMed
-
- Ferreira V, van Dijk KW, Groen AK, Vos RM, van der Kaa J, Gijbels MJ, Havekes LM, Pannekoek H. Macrophage-specific inhibition of NF-kappaB activation reduces foam-cell formation. Atherosclerosis. 2007;192:283–290. - PubMed
-
- Genet C, Strehle A, Schmidt C, Boudjelal G, Lobstein A, Schoonjans K, Souchet M, Auwerx J, Saladin R, Wagner A. Structure-activity relationship study of betulinic acid, a novel and selective TGR5 agonist, and its synthetic derivatives: potential impact in diabetes. J Med Chem. 2010;53:178–190. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
