

Modulation of phosphofructokinase (PFK) from Setaria cervi, a bovine filarial parasite, by different effectors and its interaction with some antifilarials
- PMID: 22152593
- PMCID: PMC3261128
- DOI: 10.1186/1756-3305-4-227
Modulation of phosphofructokinase (PFK) from Setaria cervi, a bovine filarial parasite, by different effectors and its interaction with some antifilarials
Abstract
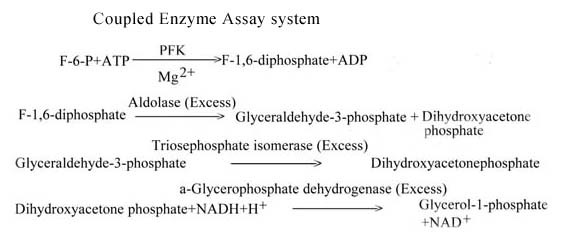
Background: Phosphofructokinase (ATP: D-fructose-6-phosphate-1-phosphotransferase, EC 2.7.1.11, PFK) is of primary importance in the regulation of glycolytic flux. This enzyme has been extensively studied from mammalian sources but relatively less attention has been paid towards its characterization from filarial parasites. Furthermore, the information about the response of filarial PFK towards the anthelmintics/antifilarial compounds is lacking. In view of these facts, PFK from Setaria cervi, a bovine filarial parasite having similarity with that of human filarial worms, was isolated, purified and characterized.
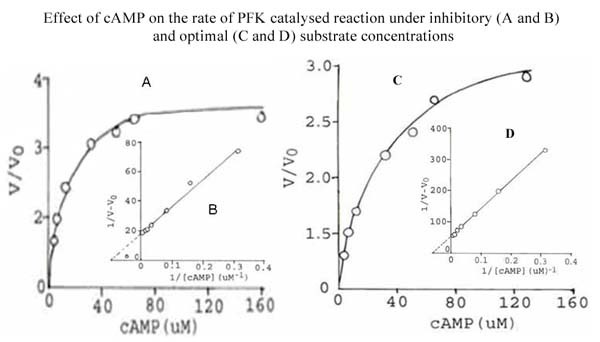
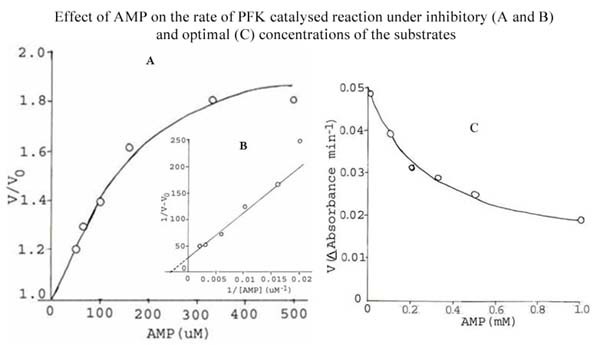
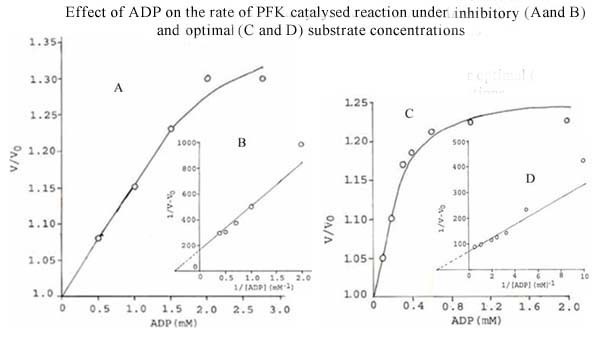
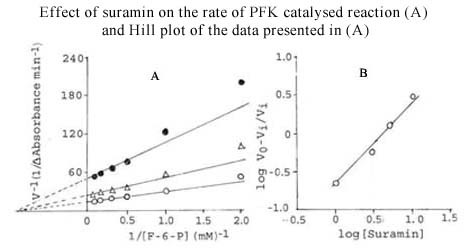
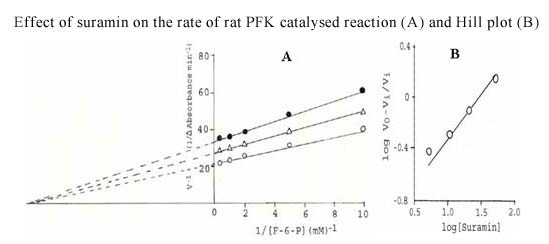
Results: The S. cervi PFK was cytosolic in nature. The adult parasites (both female and male) contained more enzyme activity than the microfilarial (Mf) stage of S. cervi, which exhibited only 20% of total activity. The S. cervi PFK could be modulated by different nucleotides and the response of enzyme to these nucleotides was dependent on the concentrations of substrates (F-6-P and ATP). The enzyme possessed wide specificity towards utilization of the nucleotides as phosphate group donors. S. cervi PFK showed the presence of thiol group(s) at the active site of the enzyme, which could be protected from inhibitory action of para-chloromercuribenzoate (p-CMB) up to about 76% by pretreatment with cysteine or β-ME. The sensitivity of PFK from S. cervi towards antifilarials/anthelmintics was comparatively higher than that of mammalian PFK. With suramin, the Ki value for rat liver PFK was 40 times higher than PFK from S. cervi.
Conclusions: The results indicate that the activity of filarial PFK may be modified by different effectors (such as nucleotides, thiol group reactants and anthelmintics) in filarial worms depending on the presence of varying concentrations of substrates (F-6-P and ATP) in the cellular milieu. It may possess thiol group at its active site responsible for catalysis. Relatively, 40 times higher sensitivity of filarial PFK towards suramin as compared to the analogous enzyme from the mammalian system indicates that this enzyme could be exploited as a potential chemotherapeutic target against filariasis.
Figures
Similar articles
-
Setaria cervi: enzymes in microfilariae and in vitro action of antifilarials.Z Parasitenkd. 1982;68(3):331-8. doi: 10.1007/BF00927411. Z Parasitenkd. 1982. PMID: 7157943
-
Kinetic Characterisation of Phosphofructokinase Purified from Setaria cervi: A Bovine Filarial Parasite.Enzyme Res. 2011;2011:939472. doi: 10.4061/2011/939472. Epub 2011 Sep 15. Enzyme Res. 2011. PMID: 21941634 Free PMC article.
-
Purification and characterization of a leucine aminopeptidase from the bovine filarial parasite Setaria cervi.Acta Trop. 2008 Apr;106(1):1-8. doi: 10.1016/j.actatropica.2007.12.009. Epub 2008 Jan 3. Acta Trop. 2008. PMID: 18262499
-
Biochemical composition and metabolic pathways of filarial worms Setaria cervi: search for new antifilarial agents.J Helminthol. 2007 Sep;81(3):261-80. doi: 10.1017/S0022149X07799133. J Helminthol. 2007. PMID: 17875226 Review.
-
Antifilarials and their mode of action.Ciba Found Symp. 1987;127:246-64. doi: 10.1002/9780470513446.ch17. Ciba Found Symp. 1987. PMID: 3297557 Review.
Cited by
-
Analysis of phosphofructokinase-1 activity as affected by pH and ATP concentration.Sci Rep. 2024 Sep 11;14(1):21192. doi: 10.1038/s41598-024-72028-4. Sci Rep. 2024. PMID: 39261563 Free PMC article.
-
Identification of ML251, a Potent Inhibitor of T. brucei and T. cruzi Phosphofructokinase.ACS Med Chem Lett. 2013 Oct 30;5(1):12-7. doi: 10.1021/ml400259d. eCollection 2014 Jan 9. ACS Med Chem Lett. 2013. PMID: 24900769 Free PMC article.
-
Enhancement of rapamycin production by metabolic engineering in Streptomyces hygroscopicus based on genome-scale metabolic model.J Ind Microbiol Biotechnol. 2017 Feb;44(2):259-270. doi: 10.1007/s10295-016-1880-1. Epub 2016 Dec 1. J Ind Microbiol Biotechnol. 2017. PMID: 27909940
References
-
- Ramachandran CP, Sivanandan S. Inoculation of infective larvae of sub-periodic Brugia malayi into domestic cats by various routes. SE Asian J Trop Med Pub Hlth. 1970;1:150. - PubMed
-
- Sharma B, Ghatak S, Malhotra OP, Kaushal NA. Stabilization and characterization of phosphofructokinase purified from Setaria cervi, a filarial bovine filarial parasite. Helminthologia. 1995;32:15–23.
-
- Sharma B. Phosphofructokinase from Setaria cervi: mode of action of certain anthelmintics /chemotherapeutics. Helminthologia. 1998;35:12–14.
-
- Bloxham DP, Lardy HA. In: The enzymes. Boyer PD, editor. Vol. 8. Academic Press, San Diego; 1973. Phosphofructokinase; p. 339.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
