

The secret languages of coevolved symbioses: insights from the Euprymna scolopes-Vibrio fischeri symbiosis
- PMID: 22154556
- PMCID: PMC3288948
- DOI: 10.1016/j.smim.2011.11.006
The secret languages of coevolved symbioses: insights from the Euprymna scolopes-Vibrio fischeri symbiosis
Abstract
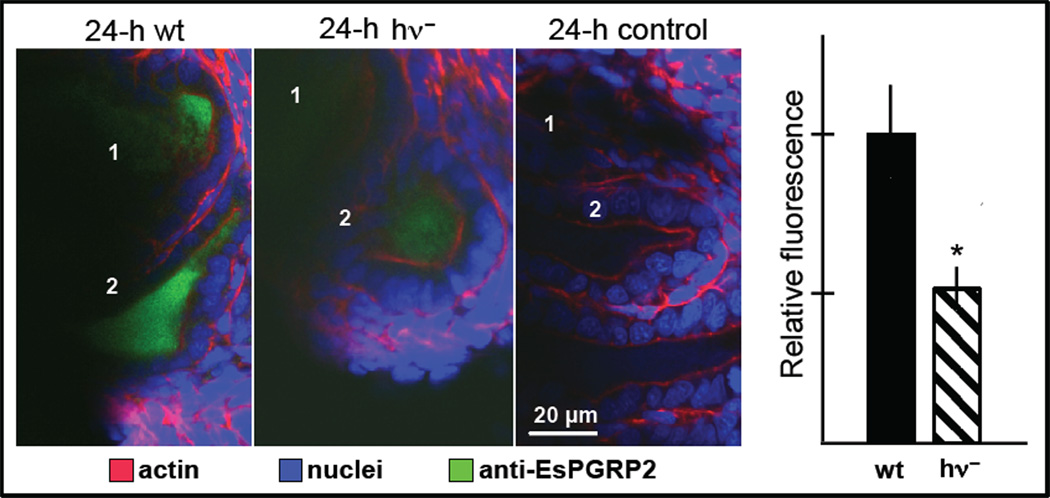
Recent research on a wide variety of systems has demonstrated that animals generally coevolve with their microbial symbionts. Although such relationships are most often established anew each generation, the partners associate with fidelity, i.e., they form exclusive alliances within the context of rich communities of non-symbiotic environmental microbes. The mechanisms by which this exclusivity is achieved and maintained remain largely unknown. Studies of the model symbiosis between the Hawaiian squid Euprymna scolopes and the marine luminous bacterium Vibrio fischeri provide evidence that the interplay between evolutionarily conserved features of the innate immune system, most notably MAMP/PRR interactions, and a specific feature of this association, i.e., luminescence, are critical for development and maintenance of this association. As such, in this partnership and perhaps others, symbiotic exclusivity is mediated by the synergism between a general animal-microbe 'language' and a 'secret language' that is decipherable only by the specific partners involved.
Copyright © 2011 Elsevier Ltd. All rights reserved.
Figures
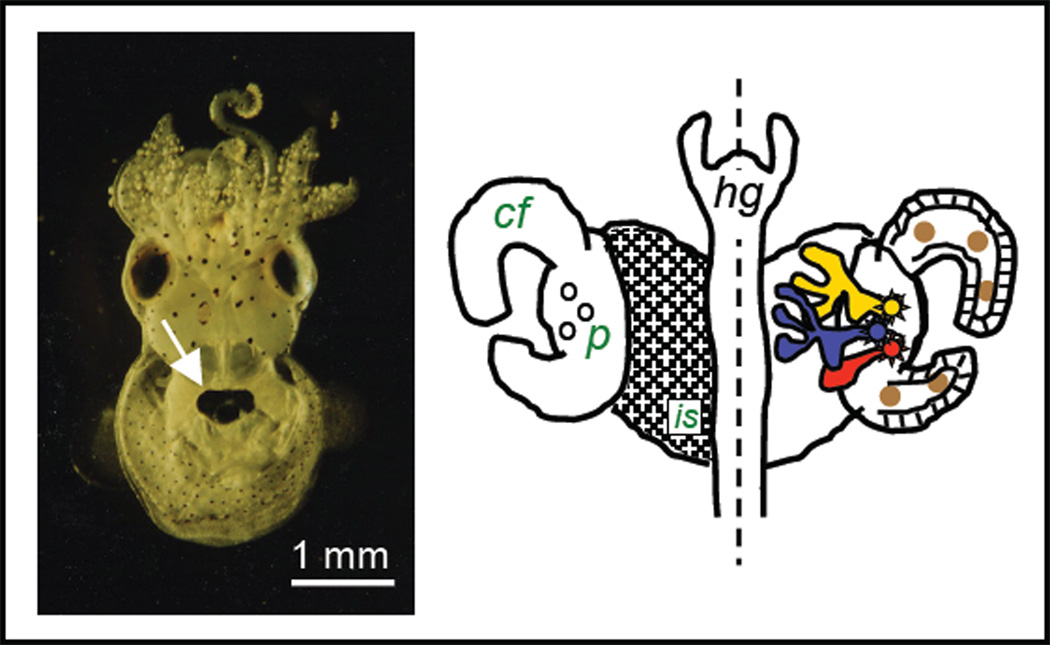
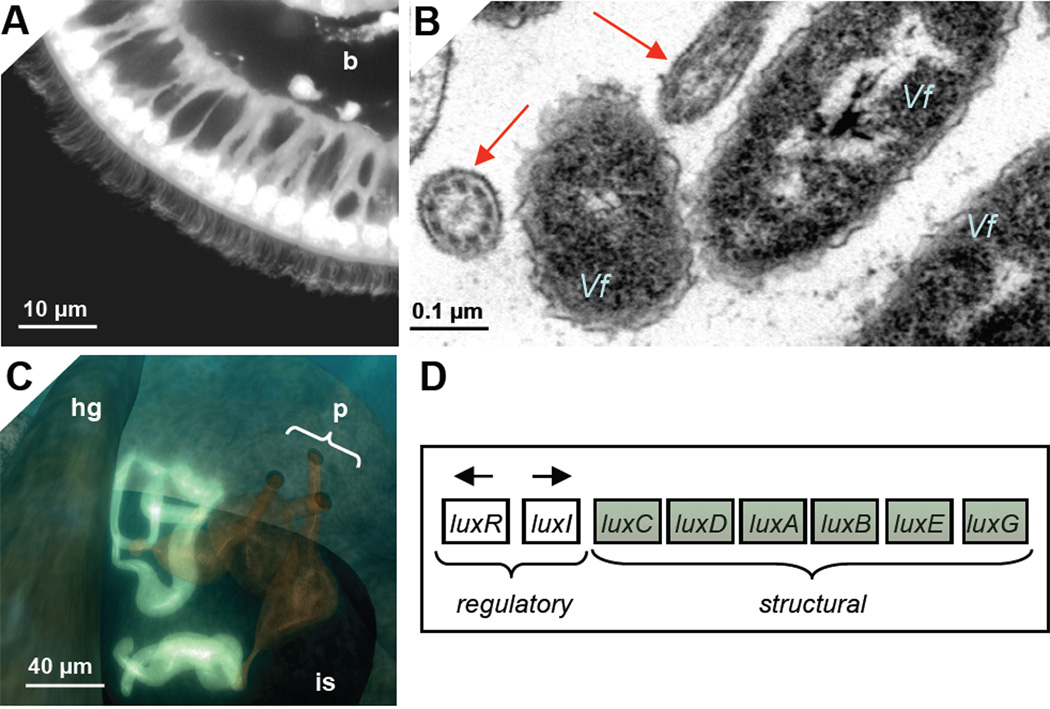
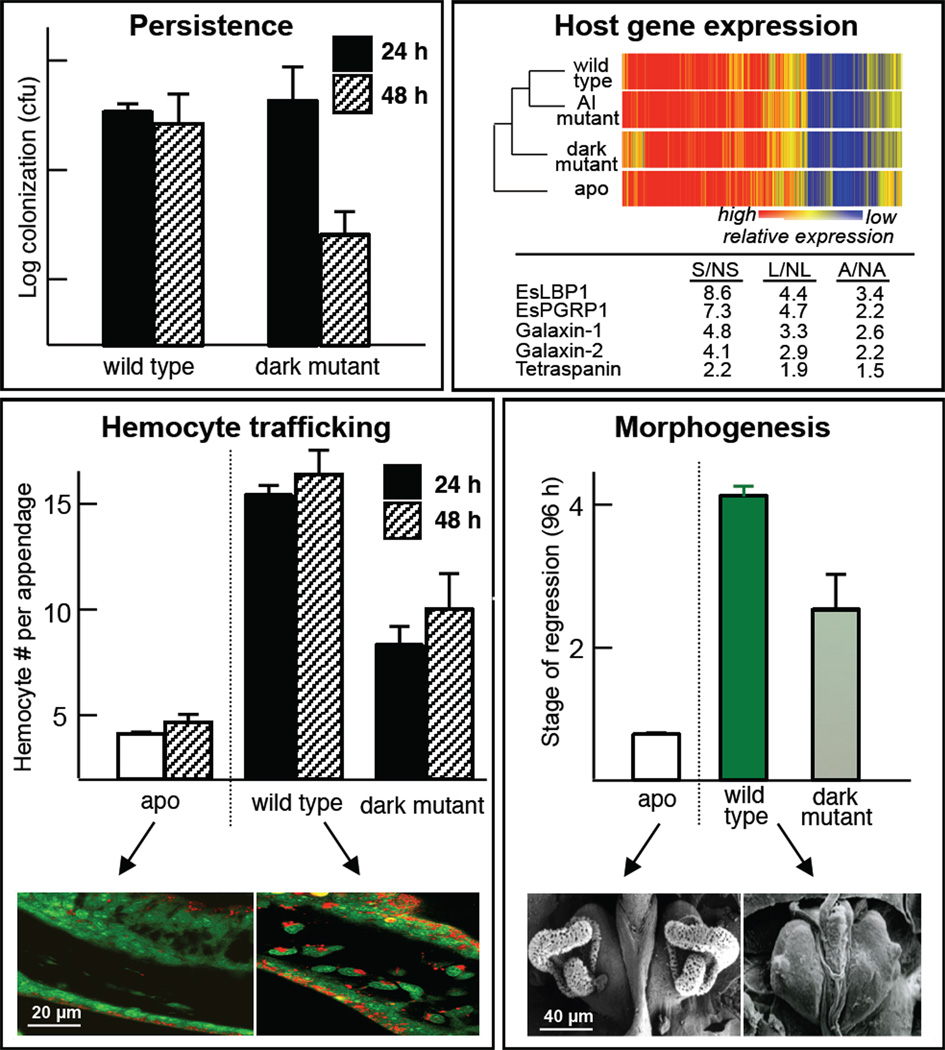
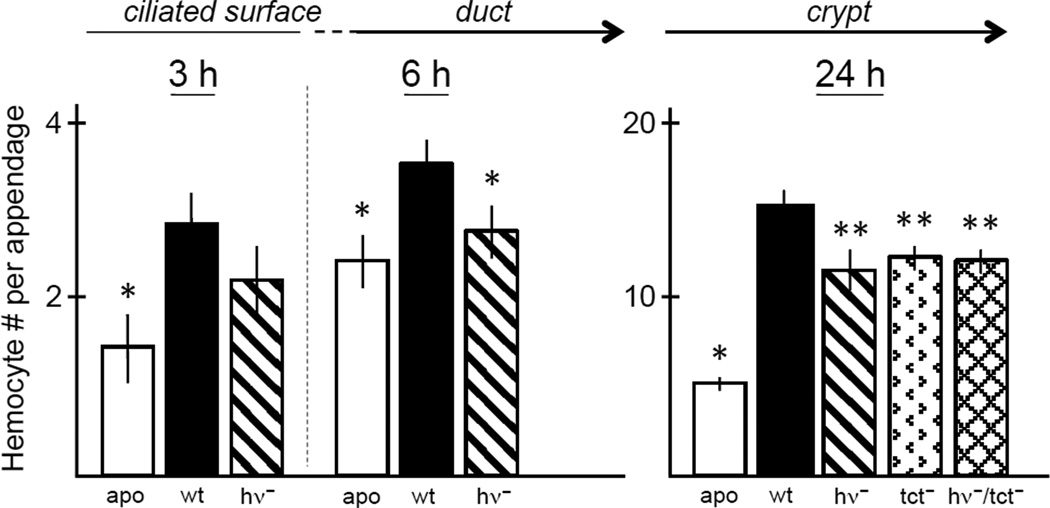
References
-
- Eberl G. A new vision of immunity: homeostasis of the superorganism. Mucosal Immunol. 2010;3:450–460. - PubMed
-
- MacDonald SJ, Thomas GH, Douglas AE. Genetic and metabolic determinants of nutritional phenotype in an insect-bacterial symbiosis. Mol Ecol. 2011;20:2073–2084. - PubMed
-
- Koropatnick TA, Engle JT, Apicella MA, Stabb EV, Goldman WE, McFall-Ngai MJ. Microbial factor-mediated development in a host-bacterial mutualism. Science. 2004;306:1186–1188. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
