

Protein-mediated protection as the predominant mechanism for defining processed mRNA termini in land plant chloroplasts
- PMID: 22156165
- PMCID: PMC3326301
- DOI: 10.1093/nar/gkr1137
Protein-mediated protection as the predominant mechanism for defining processed mRNA termini in land plant chloroplasts
Abstract
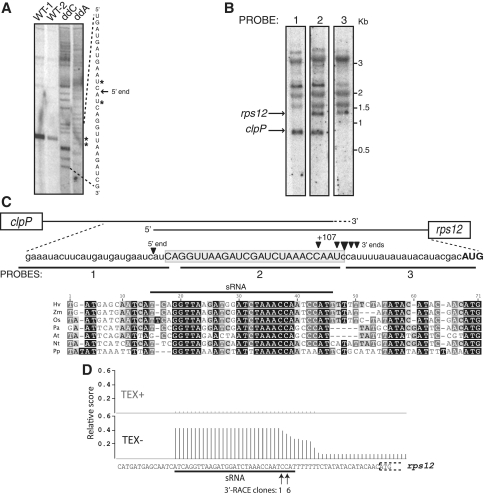
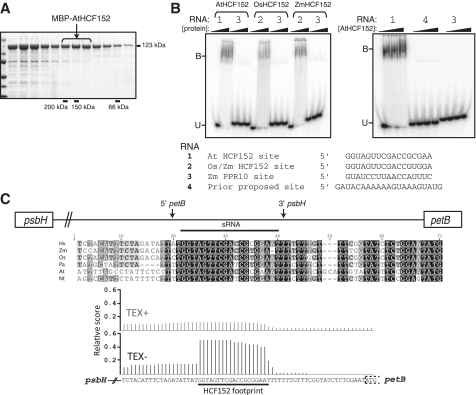
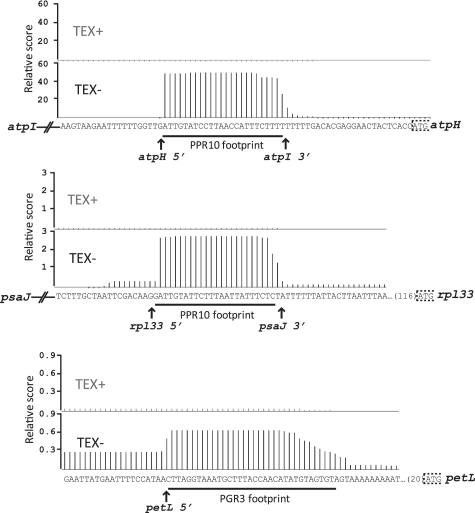
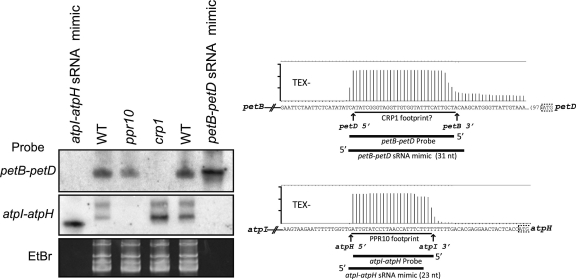
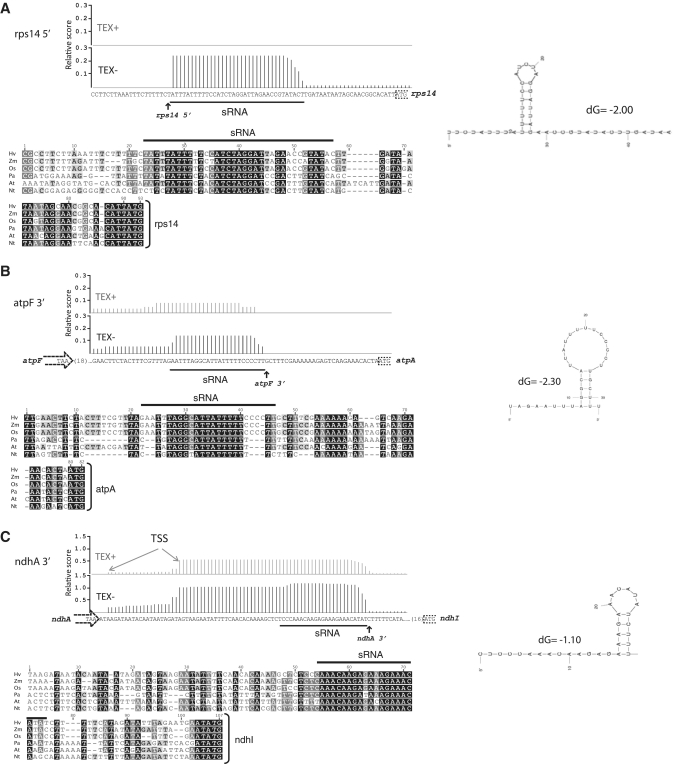
Most chloroplast mRNAs are processed from larger precursors. Several mechanisms have been proposed to mediate these processing events, including site-specific cleavage and the stalling of exonucleases by RNA structures. A protein barrier mechanism was proposed based on analysis of the pentatricopeptide repeat (PPR) protein PPR10: PPR10 binds two intercistronic regions and impedes 5'- and 3'-exonucleases, resulting in processed RNAs with PPR10 bound at the 5'- or 3'-end. In this study, we provide evidence that protein barriers are the predominant means for defining processed mRNA termini in chloroplasts. First, we map additional RNA termini whose arrangement suggests biogenesis via a PPR10-like mechanism. Second, we show that the PPR protein HCF152 binds to the immediate 5'- or 3'-termini of transcripts that require HCF152 for their accumulation, providing evidence that HCF152 defines RNA termini by blocking exonucleases. Finally, we build on the observation that the PPR10 and HCF152 binding sites accumulate as small chloroplast RNAs to infer binding sites of other PPR proteins. We show that most processed mRNA termini are represented by small RNAs whose sequences are highly conserved. We suggest that each such small RNA is the footprint of a PPR-like protein that protects the adjacent RNA from degradation.
Figures
Comment on
- Nucleic Acids Res.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
