

Nascent-seq indicates widespread cotranscriptional pre-mRNA splicing in Drosophila
- PMID: 22156210
- PMCID: PMC3243060
- DOI: 10.1101/gad.178962.111
Nascent-seq indicates widespread cotranscriptional pre-mRNA splicing in Drosophila
Abstract
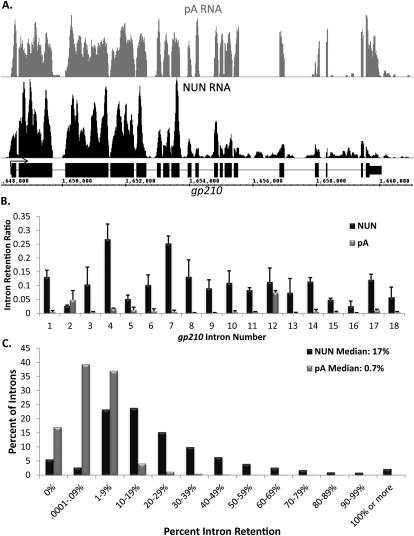
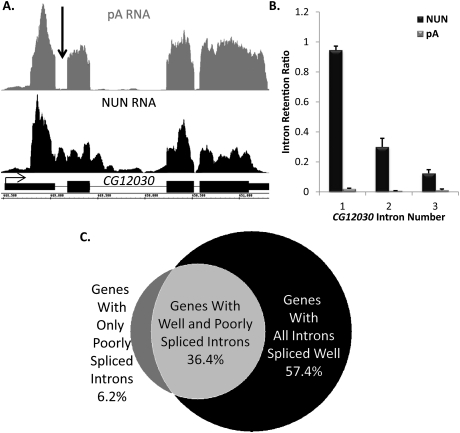
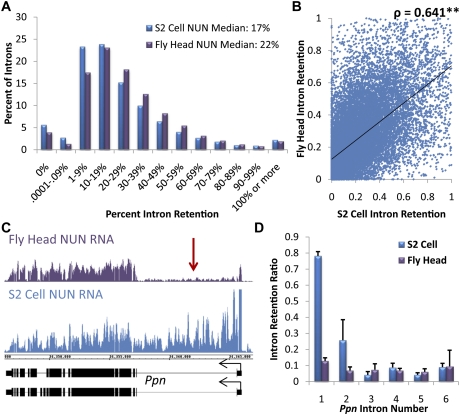
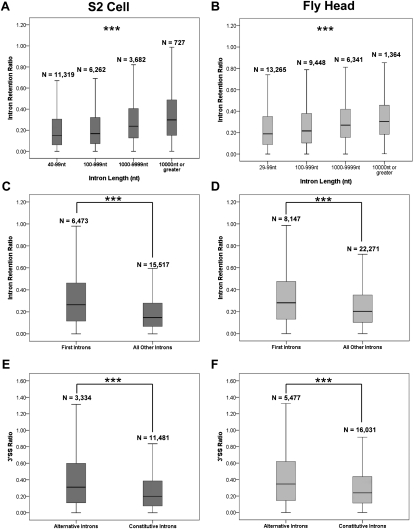
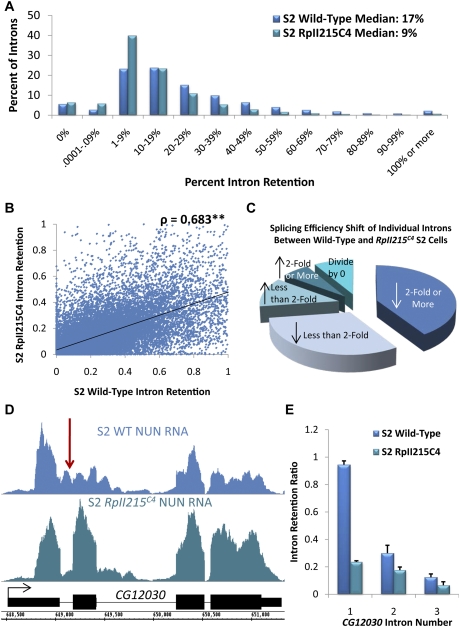
To determine the prevalence of cotranscriptional splicing in Drosophila, we sequenced nascent RNA transcripts from Drosophila S2 cells as well as from Drosophila heads. Eighty-seven percent of the introns assayed manifest >50% cotranscriptional splicing. The remaining 13% are cotranscriptionally spliced poorly or slowly, with ∼3% being almost completely retained in nascent pre-mRNA. Although individual introns showed slight but statistically significant differences in splicing efficiency, similar global levels of splicing were seen from both sources. Importantly, introns with low cotranscriptional splicing efficiencies are present in the same primary transcript with efficiently spliced introns, indicating that splicing is intron-specific. The analysis also indicates that cotranscriptional splicing is less efficient for first introns, longer introns, and introns annotated as alternative. Finally, S2 cells expressing the slow RpII215(C4) mutant show substantially less intron retention than wild-type S2 cells.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous