

Core promoter-selective function of HMGA1 and Mediator in Initiator-dependent transcription
- PMID: 22156211
- PMCID: PMC3243061
- DOI: 10.1101/gad.177360.111
Core promoter-selective function of HMGA1 and Mediator in Initiator-dependent transcription
Abstract
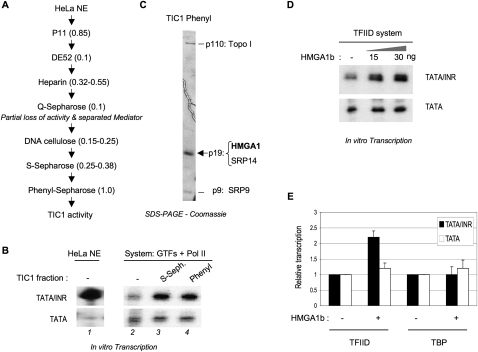
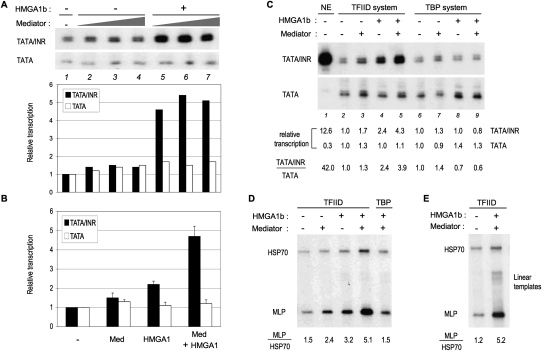
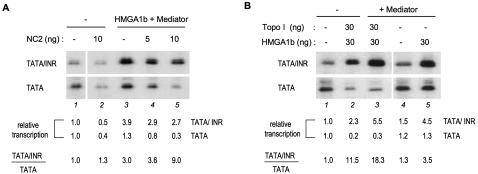
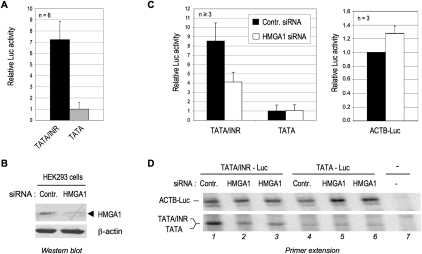
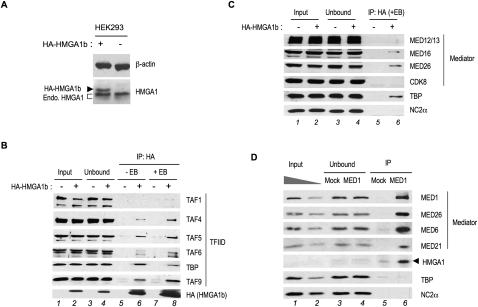
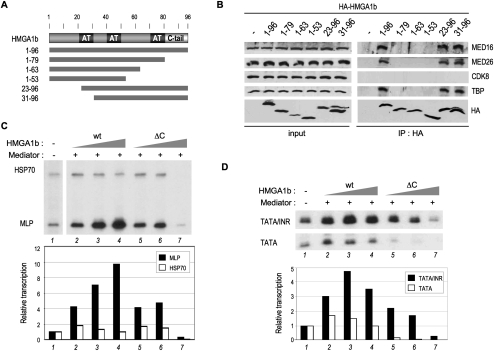
The factors and mechanisms underlying the differential activity and regulation of eukaryotic RNA polymerase II on different types of core promoters have remained elusive. Here we show that the architectural factor HMGA1 and the Mediator coregulator complex cooperate to enhance basal transcription from core promoters containing both a TATA box and an Initiator (INR) element but not from "TATA-only" core promoters. INR-dependent activation by HMGA1 and Mediator requires the TATA-binding protein (TBP)-associated factors (TAFs) within the TFIID complex and counteracts negative regulators of TBP/TATA-dependent transcription such as NC2 and Topoisomerase I. HMGA1 interacts with TFIID and Mediator and is required for the synergy of TATA and INR elements in mammalian cells. Accordingly, natural HMGA1-activated genes in embryonic stem cells tend to have both TATA and INR elements in a synergistic configuration. Our results suggest a core promoter-specific regulation of Mediator and the basal transcription machinery by HMGA1.
Figures
References
-
- Burke TW, Kadonaga JT 1996. Drosophila TFIID binds to a conserved downstream basal promoter element that is present in many TATA-box-deficient promoters. Genes Dev 10: 711–724 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources