

Multiple layers of CD80/86-dependent costimulatory activity regulate primary, memory, and secondary lymphocytic choriomeningitis virus-specific T cell immunity
- PMID: 22156513
- PMCID: PMC3302393
- DOI: 10.1128/JVI.05949-11
Multiple layers of CD80/86-dependent costimulatory activity regulate primary, memory, and secondary lymphocytic choriomeningitis virus-specific T cell immunity
Abstract
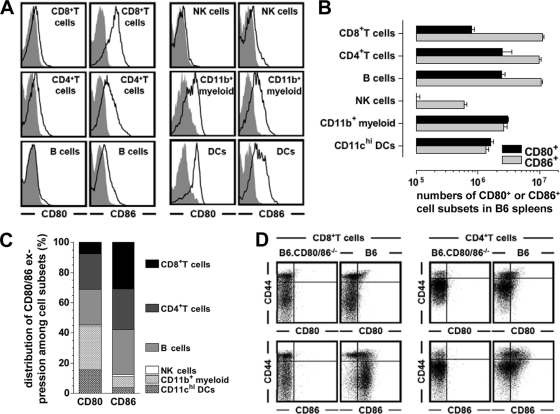
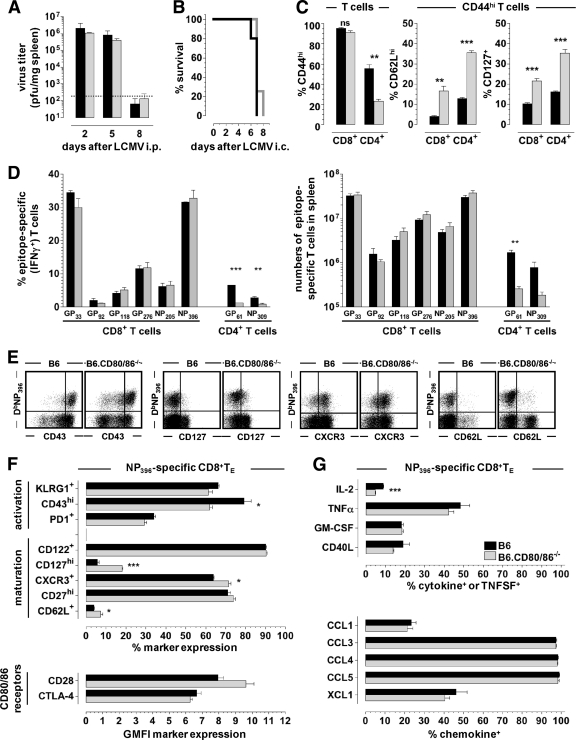
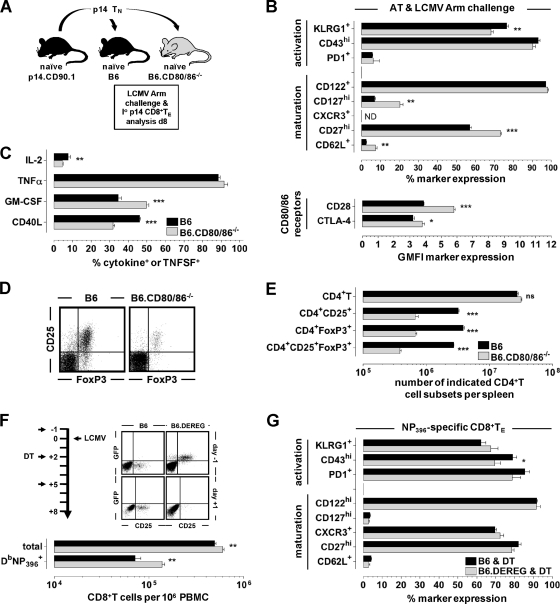
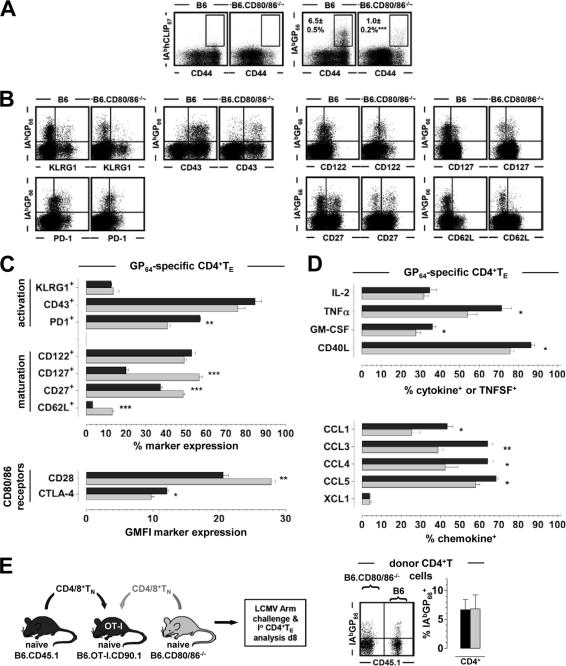
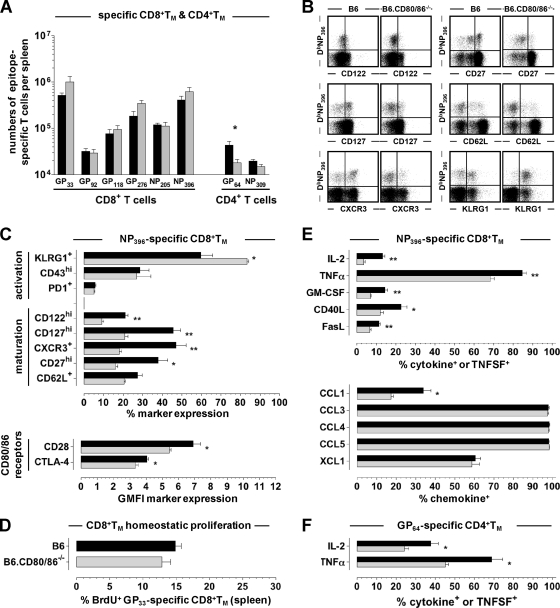
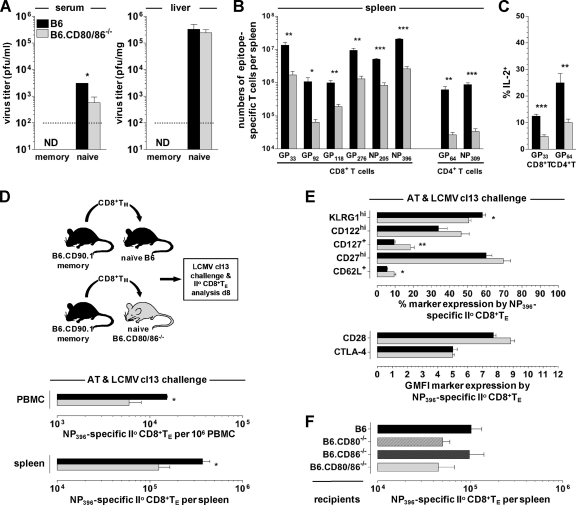
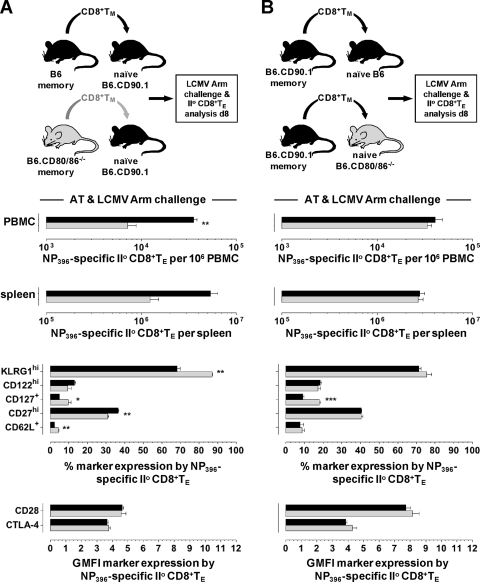
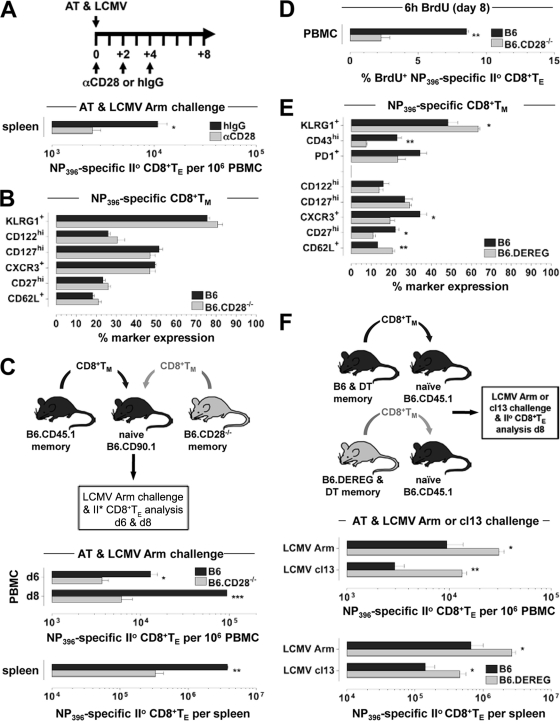
The lymphocytic choriomeningitis virus (LCMV) system constitutes one of the most widely used models for the study of infectious disease and the regulation of virus-specific T cell immunity. However, with respect to the activity of costimulatory and associated regulatory pathways, LCMV-specific T cell responses have long been regarded as relatively independent and thus distinct from the regulation of T cell immunity directed against many other viral pathogens. Here, we have reevaluated the contribution of CD28-CD80/86 costimulation in the LCMV system by use of CD80/86-deficient mice, and our results demonstrate that a disruption of CD28-CD80/86 signaling compromises the magnitude, phenotype, and/or functionality of LCMV-specific CD8(+) and/or CD4(+) T cell populations in all stages of the T cell response. Notably, a profound inhibition of secondary T cell immunity in LCMV-immune CD80/86-deficient mice emerged as a composite of both defective memory T cell development and a specific requirement for CD80 but not CD86 in the recall response, while a related experimental scenario of CD28-dependent yet CD80/86-independent secondary CD8(+) T cell immunity suggests the existence of a CD28 ligand other than CD80/86. Furthermore, we provide evidence that regulatory T cells (T(REG)s), the homeostasis of which is altered in CD80/86(-/-) mice, contribute to restrained LCMV-specific CD8(+) T cell responses in the presence of CD80/86. Our observations can therefore provide a more coherent perspective on CD28-CD80/86 costimulation in antiviral T cell immunity that positions the LCMV system within a shared context of multiple defects that virus-specific T cells acquire in the absence of CD28-CD80/86 costimulation.
Figures
Similar articles
-
The role of CD80/CD86 in generation and maintenance of functional virus-specific CD8+ T cells in mice infected with lymphocytic choriomeningitis virus.J Immunol. 2010 Aug 1;185(3):1730-43. doi: 10.4049/jimmunol.0903894. Epub 2010 Jul 2. J Immunol. 2010. PMID: 20601595
-
Role of CD28-B7 interactions in generation and maintenance of CD8 T cell memory.J Immunol. 2001 Nov 15;167(10):5565-73. doi: 10.4049/jimmunol.167.10.5565. J Immunol. 2001. PMID: 11698427
-
Role of CD40 ligand and CD28 in induction and maintenance of antiviral CD8+ effector T cell responses.J Immunol. 2000 Apr 1;164(7):3689-97. doi: 10.4049/jimmunol.164.7.3689. J Immunol. 2000. PMID: 10725727
-
Chronic LCMV Infection Is Fortified with Versatile Tactics to Suppress Host T Cell Immunity and Establish Viral Persistence.Viruses. 2021 Sep 29;13(10):1951. doi: 10.3390/v13101951. Viruses. 2021. PMID: 34696381 Free PMC article. Review.
-
Lymphocytic choriomeningitis infection of the central nervous system.Front Biosci. 2008 May 1;13:4529-43. doi: 10.2741/3021. Front Biosci. 2008. PMID: 18508527 Free PMC article. Review.
Cited by
-
In vivo identification of apoptotic and extracellular vesicle-bound live cells using image-based deep learning.J Extracell Vesicles. 2020 Jul 16;9(1):1792683. doi: 10.1080/20013078.2020.1792683. J Extracell Vesicles. 2020. PMID: 32944180 Free PMC article.
-
The viral context instructs the redundancy of costimulatory pathways in driving CD8(+) T cell expansion.Elife. 2015 Aug 11;4:e07486. doi: 10.7554/eLife.07486. Elife. 2015. PMID: 26263500 Free PMC article.
-
Extrinsic Acquisition of CD80 by Antigen-Specific CD8+ T Cells Regulates Their Recall Immune Responses to Acute Viral Infection.Immune Netw. 2019 Aug 16;19(4):e25. doi: 10.4110/in.2019.19.e25. eCollection 2019 Aug. Immune Netw. 2019. PMID: 31501713 Free PMC article.
-
Rescue of exhausted CD8 T cells by PD-1-targeted therapies is CD28-dependent.Science. 2017 Mar 31;355(6332):1423-1427. doi: 10.1126/science.aaf0683. Epub 2017 Mar 9. Science. 2017. PMID: 28280249 Free PMC article.
-
Chemokine Signatures of Pathogen-Specific T Cells II: Memory T Cells in Acute and Chronic Infection.J Immunol. 2020 Oct 15;205(8):2188-2206. doi: 10.4049/jimmunol.2000254. Epub 2020 Sep 18. J Immunol. 2020. PMID: 32948682 Free PMC article.
References
-
- Andreasen SO, Christensen JE, Marker O, Thomsen AR. 2000. Role of CD40 ligand and CD28 in induction and maintenance of antiviral CD8+ effector T cell responses. J. Immunol. 164:3689–3697 - PubMed
-
- Bertram EM, et al. 2004. A switch in costimulation from CD28 to 4-1BB during primary versus secondary CD8 T cell response to influenza in vivo. J. Immunol. 172:981–988 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
