

Toward convergence of experimental studies and theoretical modeling of the chromatin fiber
- PMID: 22157002
- PMCID: PMC3285299
- DOI: 10.1074/jbc.R111.305763
Toward convergence of experimental studies and theoretical modeling of the chromatin fiber
Abstract
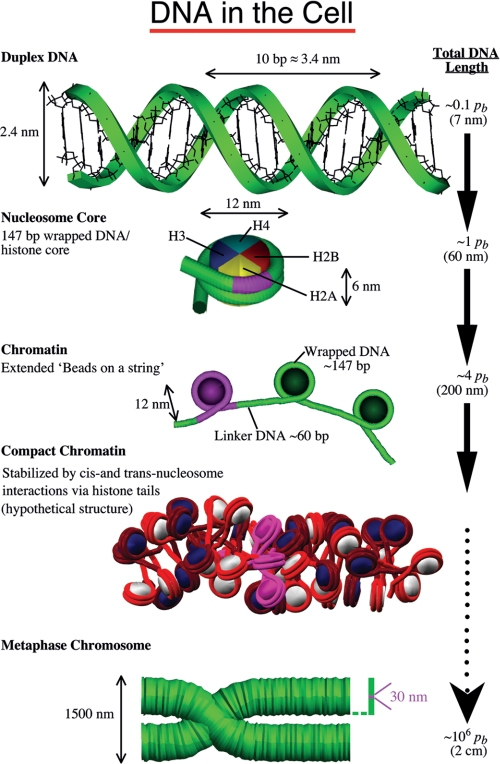
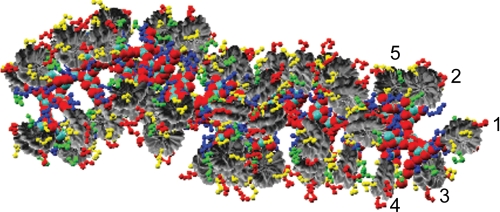
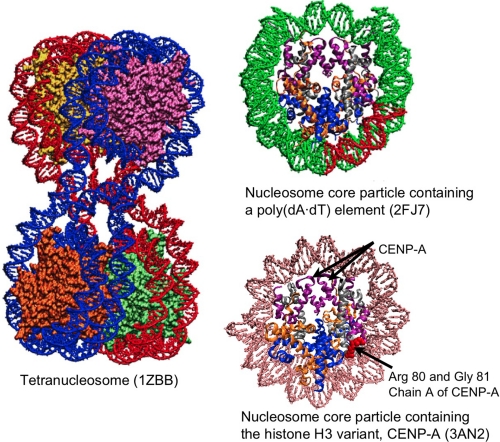
Understanding the structural organization of eukaryotic chromatin and its control of gene expression represents one of the most fundamental and open challenges in modern biology. Recent experimental advances have revealed important characteristics of chromatin in response to changes in external conditions and histone composition, such as the conformational complexity of linker DNA and histone tail domains upon compact folding of the fiber. In addition, modeling studies based on high-resolution nucleosome models have helped explain the conformational features of chromatin structural elements and their interactions in terms of chromatin fiber models. This minireview discusses recent progress and evidence supporting structural heterogeneity in chromatin fibers, reconciling apparently contradictory fiber models.
Figures


References
-
- Woodcock C. L., Dimitrov S. (2001) Higher order structure of chromatin and chromosomes. Curr. Opin. Genet. Dev. 11, 130–135 - PubMed
-
- van Holde K. E. (1988) Chromatin, Springer-Verlag, New York
-
- Felsenfeld G., Groudine M. (2003) Controlling the double helix. Nature 421, 448–453 - PubMed
-
- Horn P. J., Peterson C. L. (2002) Molecular biology. Chromatin higher order folding–wrapping up transcription. Science 297, 1824–1827 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
