

Review
doi: 10.1074/jbc.R111.275578.
Epub 2011 Dec 6.
Analysis and functional prediction of reactive cysteine residues
Affiliations
- PMID: 22157013
- PMCID: PMC3281665
- DOI: 10.1074/jbc.R111.275578
Item in Clipboard
Review
Analysis and functional prediction of reactive cysteine residues
J Biol Chem.
.
Abstract
Cys is much different from other common amino acids in proteins. Being one of the least abundant residues, Cys is often observed in functional sites in proteins. This residue is reactive, polarizable, and redox-active; has high affinity for metals; and is particularly responsive to the local environment. A better understanding of the basic properties of Cys is essential for interpretation of high-throughput data sets and for prediction and classification of functional Cys residues. We provide an overview of approaches used to study Cys residues, from methods for investigation of their basic properties, such as exposure and pK(a), to algorithms for functional prediction of different types of Cys in proteins.
Figures
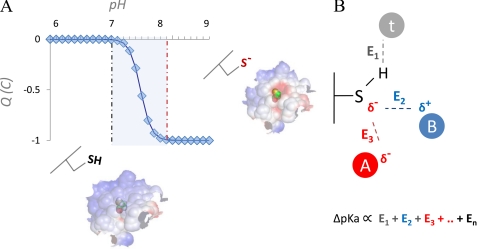
Effect of pH perturbation on net charge of exposed Cys and electrostatic properties of molecular surface. Exposed Cys residues are significantly more polar than buried Cys residues, a consequence of the high degree of Cys polarizability. For many exposed and polar Cys residues, pKa is close to the physiological pH (A, blue, shaded). In such cases, for a typical monoprotic acid in a physiological solution (assuming Henderson-Hasselbalch behavior), sudden negative charge switches can occur in the response to even very limited local pH shifts (A, neutral Cys for lower pH values and anionic Cys for higher pH values; note the steep transition between the two Cys forms in the shaded area). For any Cys in the protein, its interactions with other titratable groups and solvent determine the degree of reactivity of that Cys and influence its pKa. From a computational perspective, a common way to quantify the pKa of a residue is to calculate its deviation (ΔpKa) from the reference pKa value (pKa(REF)) for that amino acid type; ΔpKa is derived by properly accounting for all interactions with the titratable functional group, e.g. E1, E2, … En in B, where interactions with the generic titratable residues A (red circle; indicating an interacting acidic residue), B (blue circle, indicating an interacting basic residue), and t (gray circle; indicating a generic non-charged titratable residue, e.g. Thr) are shown. The ΔpKa then allows the pKa of a residue to be expressed as pKa = pKa(REF) + ΔpKa.
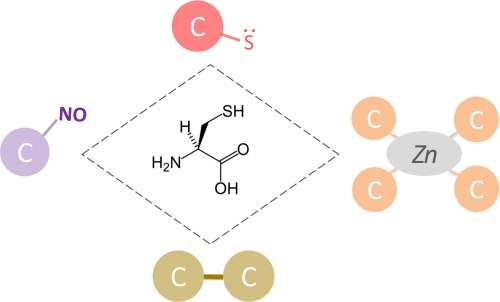
Different functional categories of Cys residues. A schematic representation of Cys functionality is shown. Starting from the top of the rhombus, which gives the molecular structure of Cys, and going clockwise: catalytic residues (red circle; representing a nucleophilic thiolate), metal-binding Cys (orange circles; representing a zinc-Cys4-binding site), structural disulfides (yellow circles; representing a covalent bond between two Cys residues), and regulatory Cys (violet circle; representing an S-nitrosylation site). As discussed in text, not all functional Cys can be reliably categorized. For example, some catalytic Cys residues can also be S-nitrosylated or oxidized to sulfenic acid, and some metal-binding Cys residues can, in certain situations, turn to cystine residues. To represent this complexity and an occasional interplay of functions, the rhombus connecting different functional Cys categories is shown with a dashed line.
References
-
- Trifonov E. N. (2004) The triplet code from first principles. J. Biomol. Struct. Dyn. 22, 1–11 - PubMed
-
- Jordan I. K., Kondrashov F. A., Adzhubei I. A., Wolf Y. I., Koonin E. V., Kondrashov A. S., Sunyaev S. (2005) A universal trend of amino acid gain and loss in protein evolution. Nature 433, 633–638 - PubMed
-
- Wu H., Ma B. G., Zhao J. T., Zhang H. Y. (2007) How similar are amino acid mutations in human genetic diseases and evolution. Biochem. Biophys. Res. Commun. 362, 233–237 - PubMed
-
- Moosmann B., Behl C. (2008) Mitochondrially encoded cysteine predicts animal life span. Aging Cell 7, 32–46 - PubMed
-
- Jobson R. W., Dehne-Garcia A., Galtier N. (2010) Apparent longevity-related adaptation of mitochondrial amino acid content is due to nucleotide compositional shifts. Mitochondrion 10, 540–547 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
