

Lipin-1 phosphatidic phosphatase activity modulates phosphatidate levels to promote peroxisome proliferator-activated receptor γ (PPARγ) gene expression during adipogenesis
- PMID: 22157014
- PMCID: PMC3271002
- DOI: 10.1074/jbc.M111.296681
Lipin-1 phosphatidic phosphatase activity modulates phosphatidate levels to promote peroxisome proliferator-activated receptor γ (PPARγ) gene expression during adipogenesis
Abstract
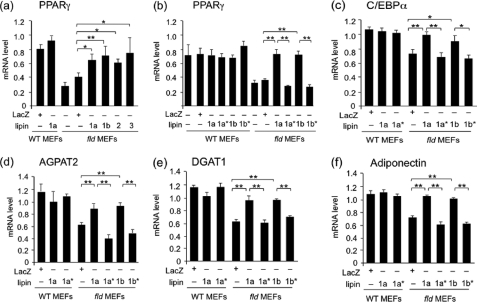
Adipose tissue plays a key role in metabolic homeostasis. Disruption of the Lpin1 gene encoding lipin-1 causes impaired adipose tissue development and function in rodents. Lipin-1 functions as a phosphatidate phosphatase (PAP) enzyme in the glycerol 3-phosphate pathway for triglyceride storage and as a transcriptional coactivator/corepressor for metabolic nuclear receptors. Previous studies established that lipin-1 is required at an early step in adipocyte differentiation for induction of the adipogenic gene transcription program, including the key regulator peroxisome proliferator-activated receptor γ (PPARγ). Here, we investigate the requirement of lipin-1 PAP versus coactivator function in the establishment of Pparg expression during adipocyte differentiation. We demonstrate that PAP activity supplied by lipin-1, lipin-2, or lipin-3, but not lipin-1 coactivator activity, can rescue Pparg gene expression and lipogenesis during adipogenesis in lipin-1-deficient preadipocytes. In adipose tissue from lipin-1-deficient mice, there is an accumulation of phosphatidate species containing a range of medium chain fatty acids and an activation of the MAPK/extracellular signal-related kinase (ERK) signaling pathway. Phosphatidate inhibits differentiation of cultured adipocytes, and this can be rescued by the expression of lipin-1 PAP activity or by inhibition of ERK signaling. These results emphasize the importance of lipid intermediates as choreographers of gene regulation during adipogenesis, and the results highlight a specific role for lipins as determinants of levels of a phosphatidic acid pool that influences Pparg expression.
Figures




References
-
- Huang-Doran I., Sleigh A., Rochford J. J., O'Rahilly S., Savage D. B. (2010) Lipodystrophy. Metabolic insights from a rare disorder. J. Endocrinol. 207, 245–255 - PubMed
-
- Reue K., Phan J. (2006) Metabolic consequences of lipodystrophy in mouse models. Curr. Opin. Clin. Nutr. Metab. Care 9, 436–441 - PubMed
-
- Zalesin K. C., Franklin B. A., Miller W. M., Peterson E. D., McCullough P. A. (2011) Impact of obesity on cardiovascular disease. Med. Clin. North Am. 95, 919–937 - PubMed
-
- Lowe C. E., O'Rahilly S., Rochford J. J. (2011) Adipogenesis at a glance. J. Cell Sci. 124, 2681–2686 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
