

Structural basis for increased toxicity of pathological aβ42:aβ40 ratios in Alzheimer disease
- PMID: 22157754
- PMCID: PMC3285338
- DOI: 10.1074/jbc.M111.264473
Structural basis for increased toxicity of pathological aβ42:aβ40 ratios in Alzheimer disease
Abstract
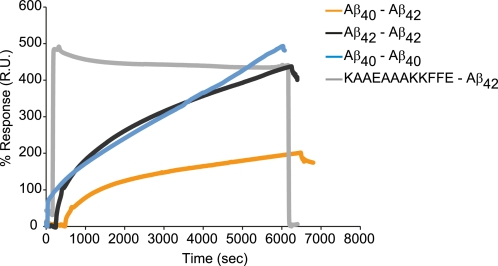
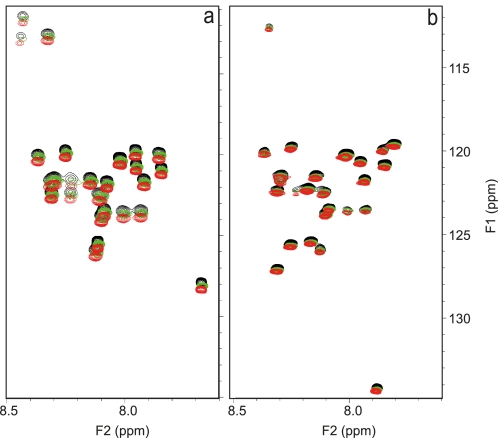
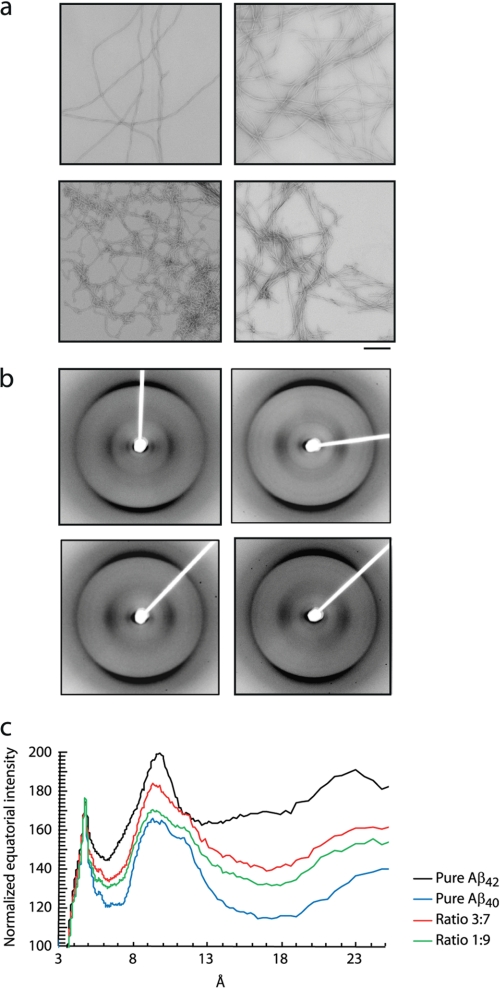
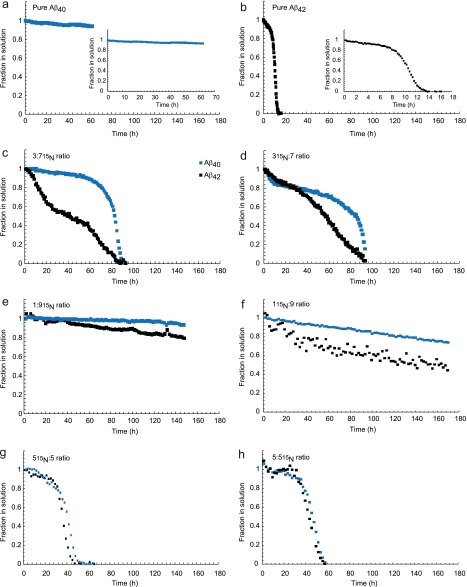
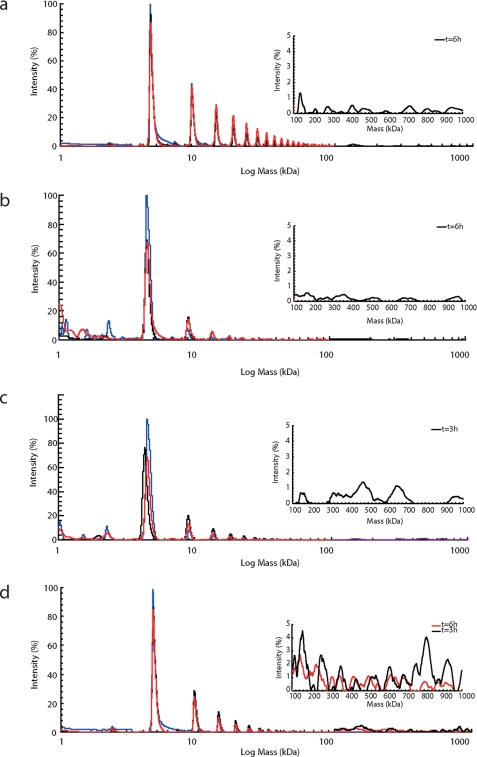
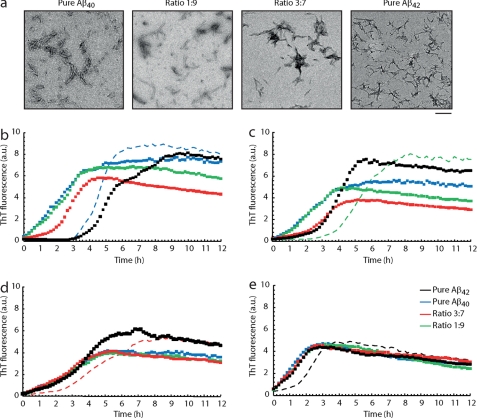
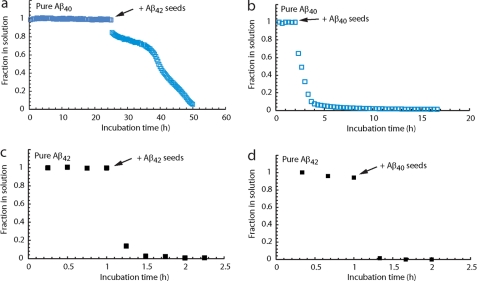
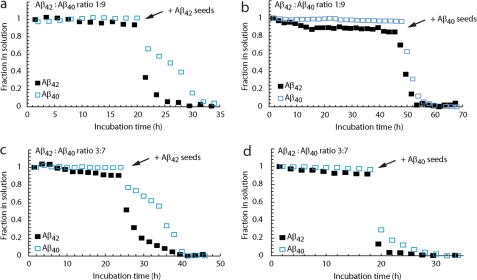
The β-amyloid peptide (Aβ) is directly related to neurotoxicity in Alzheimer disease (AD). The two most abundant alloforms of the peptide co-exist under normal physiological conditions in the brain in an Aβ(42):Aβ(40) ratio of ∼1:9. This ratio is often shifted to a higher percentage of Aβ(42) in brains of patients with familial AD and this has recently been shown to lead to increased synaptotoxicity. The molecular basis for this phenomenon is unclear. Although the aggregation characteristics of Aβ(40) and Aβ(42) individually are well established, little is known about the properties of mixtures. We have explored the biophysical and structural properties of physiologically relevant Aβ(42):Aβ(40) ratios by several techniques. We show that Aβ(40) and Aβ(42) directly interact as well as modify the behavior of the other. The structures of monomeric and fibrillar assemblies formed from Aβ(40) and Aβ(42) mixtures do not differ from those formed from either of these peptides alone. Instead, the co-assembly of Aβ(40) and Aβ(42) influences the aggregation kinetics by altering the pattern of oligomer formation as evidenced by a unique combination of solution nuclear magnetic resonance spectroscopy, high molecular weight mass spectrometry, and cross-seeding experiments. We relate these observations to the observed enhanced toxicity of relevant ratios of Aβ(42):Aβ(40) in synaptotoxicity assays and in AD patients.
Figures
References
-
- McLean C. A., Cherny R. A., Fraser F. W., Fuller S. J., Smith M. J., Beyreuther K., Bush A. I., Masters C. L. (1999) Soluble pool of Aβ amyloid as a determinant of severity of neurodegeneration in Alzheimer disease. Ann. Neurol. 46, 860–866 - PubMed
-
- Lambert M. P., Barlow A. K., Chromy B. A., Edwards C., Freed R., Liosatos M., Morgan T. E., Rozovsky I., Trommer B., Viola K. L., Wals P., Zhang C., Finch C. E., Krafft G. A., Klein W. L. (1998) Diffusible, nonfibrillar ligands derived from Aβ1–42 are potent central nervous system neurotoxins. Proc. Natl. Acad. Sci. U.S.A. 95, 6448–6453 - PMC - PubMed
-
- Klyubin I., Betts V., Welzel A. T., Blennow K., Zetterberg H., Wallin A., Lemere C. A., Cullen W. K., Peng Y., Wisniewski T., Selkoe D. J., Anwyl R., Walsh D. M., Rowan M. J. (2008) Amyloid β protein dimer-containing human CSF disrupts synaptic plasticity: prevention by systemic passive immunization. J. Neurosci. 28, 4231–4237 - PMC - PubMed
-
- Shankar G. M., Li S., Mehta T. H., Garcia-Munoz A., Shepardson N. E., Smith I., Brett F. M., Farrell M. A., Rowan M. J., Lemere C. A., Regan C. M., Walsh D. M., Sabatini B. L., Selkoe D. J. (2008) Amyloid β protein dimers isolated directly from Alzheimer brains impair synaptic plasticity and memory. Nat. Med. 14, 837–842 - PMC - PubMed
-
- Lashuel H. A., Hartley D., Petre B. M., Walz T., Lansbury P. T., Jr. (2002) Nature 418, 291. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
