

The tudor domain protein kumo is required to assemble the nuage and to generate germline piRNAs in Drosophila
- PMID: 22157814
- PMCID: PMC3280549
- DOI: 10.1038/emboj.2011.449
The tudor domain protein kumo is required to assemble the nuage and to generate germline piRNAs in Drosophila
Abstract
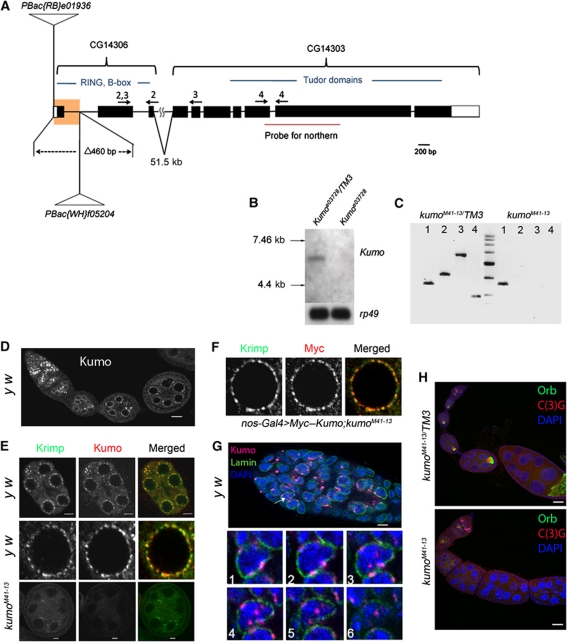
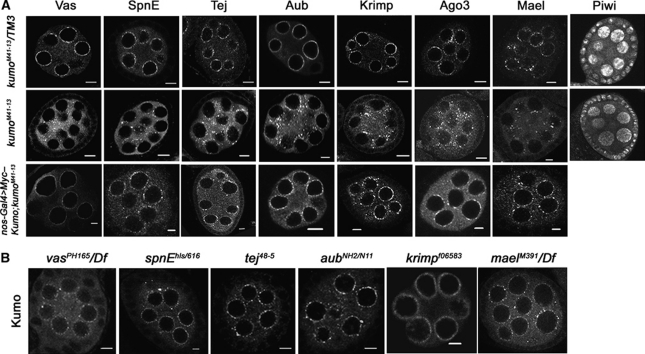
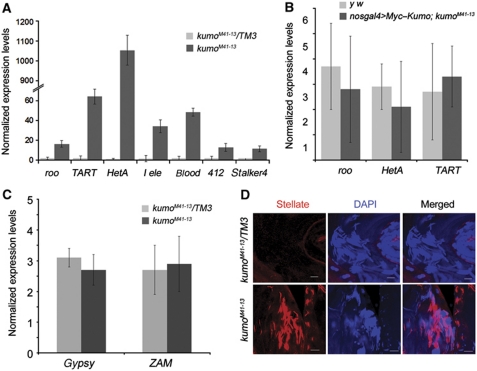
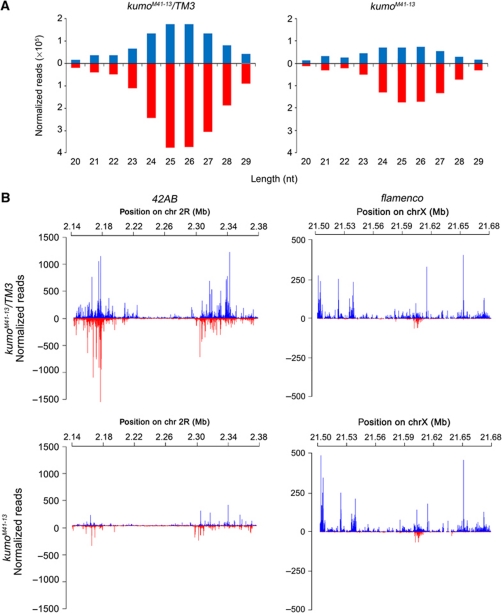
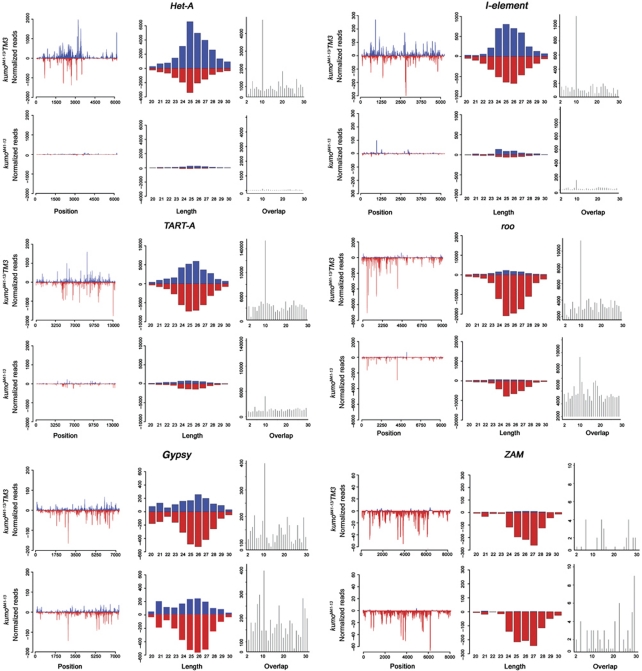
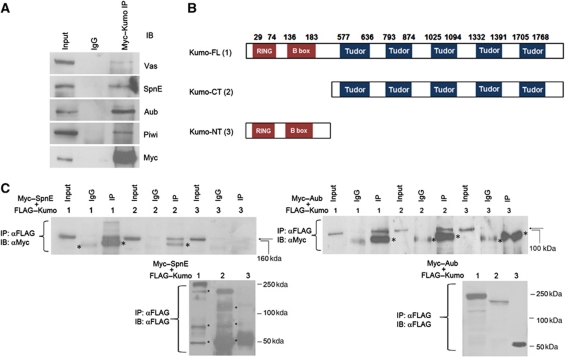
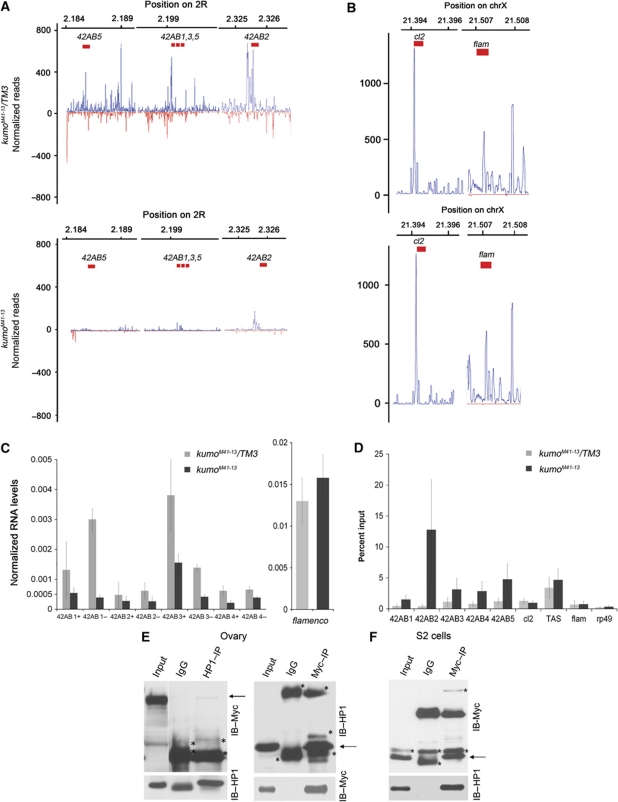
In Drosophila ovaries, distinct Piwi-interacting RNA (piRNA) pathways defend against transposons in somatic and germline cells. Germline piRNAs predominantly arise from bidirectional clusters and are amplified by the ping-pong cycle. In this study, we characterize a novel Drosophila gene, kumo and show that it encodes a conserved germline piRNA pathway component. Kumo contains five tudor domains and localizes to nuage, a unique structure present in animal germline cells, which is considered to be the processing site for germline piRNAs. Transposons targeted by the germline piRNA pathway are derepressed in kumo mutant females. Moreover, germline piRNA production is significantly reduced in mutant ovaries, thereby indicating that kumo is required to generate germline piRNAs. Kumo localizes to the nuage as well as to nucleus early female germ cells, where it is required to maintain cluster transcript levels. Our data suggest that kumo facilitates germline piRNA production by promoting piRNA cluster transcription in the nucleus and piRNA processing at the nuage.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
Comment in
-
Antisense piRNA amplification, but not piRNA production or nuage assembly, requires the Tudor-domain protein Qin.EMBO J. 2014 Mar 18;33(6):536-9. doi: 10.1002/embj.201384895. EMBO J. 2014. PMID: 24652836 Free PMC article. No abstract available.
-
Response to Zhang et al.EMBO J. 2014 Mar 18;33(6):540-1. doi: 10.1002/embj.201387548. EMBO J. 2014. PMID: 24652837 Free PMC article. No abstract available.
References
-
- Aravin AA, Naumova NM, Tulin AV, Vagin VV, Rozovsky YM, Gvozdev VA (2001) Double-stranded RNA-mediated silencing of genomic tandem repeats and transposable elements in the D. melanogaster germline. Curr Biol 11: 1017–1027 - PubMed
-
- Brennecke J, Aravin AA, Stark A, Dus M, Kellis M, Sachidanandam R, Hannon GJ (2007) Discrete small RNA-generating loci as master regulators of transposon activity in Drosophila. Cell 128: 1089–1103 - PubMed
-
- Eddy EM (1974) Fine structural observations on the form and distribution of nuage in germ cells of the rat. Anat Rec 178: 731–757 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
