

Acquisition of a multifunctional IgA+ plasma cell phenotype in the gut
- PMID: 22158124
- PMCID: PMC3487691
- DOI: 10.1038/nature10698
Acquisition of a multifunctional IgA+ plasma cell phenotype in the gut
Abstract
The largest mucosal surface in the body is in the gastrointestinal tract, a location that is heavily colonized by microbes that are normally harmless. A key mechanism required for maintaining a homeostatic balance between this microbial burden and the lymphocytes that densely populate the gastrointestinal tract is the production and transepithelial transport of poly-reactive IgA (ref. 1). Within the mucosal tissues, B cells respond to cytokines, sometimes in the absence of T-cell help, undergo class switch recombination of their immunoglobulin receptor to IgA, and differentiate to become plasma cells. However, IgA-secreting plasma cells probably have additional attributes that are needed for coping with the tremendous bacterial load in the gastrointestinal tract. Here we report that mouse IgA(+) plasma cells also produce the antimicrobial mediators tumour-necrosis factor-α (TNF-α) and inducible nitric oxide synthase (iNOS), and express many molecules that are commonly associated with monocyte/granulocytic cell types. The development of iNOS-producing IgA(+) plasma cells can be recapitulated in vitro in the presence of gut stroma, and the acquisition of this multifunctional phenotype in vivo and in vitro relies on microbial co-stimulation. Deletion of TNF-α and iNOS in B-lineage cells resulted in a reduction in IgA production, altered diversification of the gut microbiota and poor clearance of a gut-tropic pathogen. These findings reveal a novel adaptation to maintaining homeostasis in the gut, and extend the repertoire of protective responses exhibited by some B-lineage cells.
Conflict of interest statement
Figures
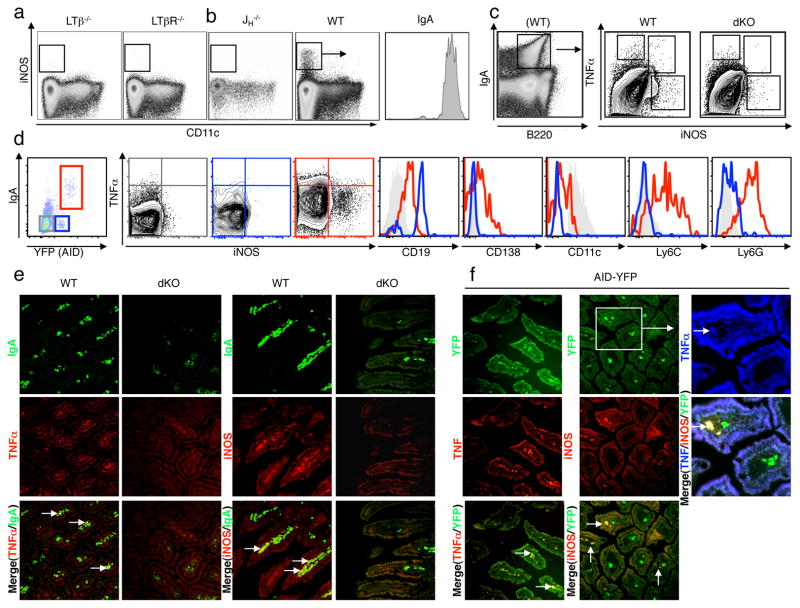
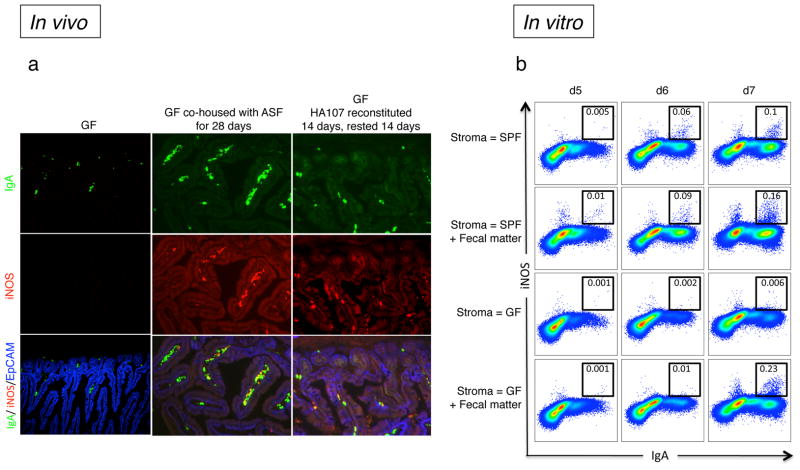
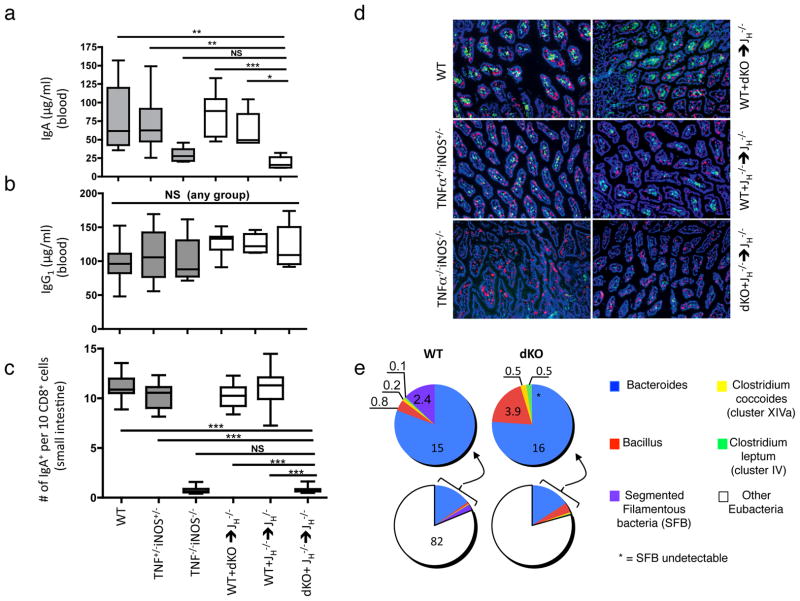
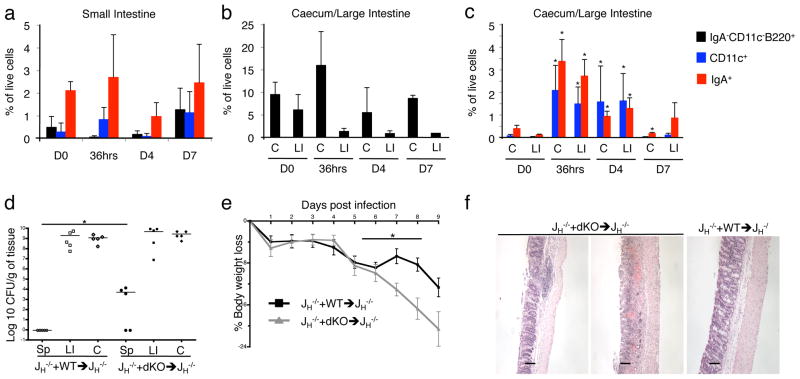
Comment in
-
Mucosal immunology: Multifunctional gut IgA⁺ plasma cells.Nat Rev Immunol. 2012 Jan 13;12(2):75. doi: 10.1038/nri3157. Nat Rev Immunol. 2012. PMID: 22240626 No abstract available.
References
-
- Hooper LV, Macpherson AJ. Immune adaptations that maintain homeostasis with the intestinal microbiota. Nat Rev Immunol. 2010;10:159–169. - PubMed
-
- Fagarasan S, Kawamoto S, Kanagawa O, Suzuki K. Adaptive immune regulation in the gut: T cell-dependent and T cell-independent IgA synthesis. Annu Rev Immunol. 2010;28:243–273. - PubMed
-
- Lee MR, Seo GY, Kim YM, Kim PH. iNOS potentiates mouse Ig isotype switching through AID expression. Biochem Biophys Res Commun. 2011 - PubMed
-
- Tezuka H, et al. Regulation of IgA production by naturally occurring TNF/iNOS-producing dendritic cells. Nature. 2007;448:929–933. - PubMed
-
- Kang HS, et al. Signaling via LTbetaR on the lamina propria stromal cells of the gut is required for IgA production. Nat Immunol. 2002;3:576–582. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
