

Temperature regulation of virulence factors in the pathogen Vibrio coralliilyticus
- PMID: 22158392
- PMCID: PMC3309362
- DOI: 10.1038/ismej.2011.154
Temperature regulation of virulence factors in the pathogen Vibrio coralliilyticus
Abstract
Sea surface temperatures (SST) are rising because of global climate change. As a result, pathogenic Vibrio species that infect humans and marine organisms during warmer summer months are of growing concern. Coral reefs, in particular, are already experiencing unprecedented degradation worldwide due in part to infectious disease outbreaks and bleaching episodes that are exacerbated by increasing SST. For example, Vibrio coralliilyticus, a globally distributed bacterium associated with multiple coral diseases, infects corals at temperatures above 27 °C. The mechanisms underlying this temperature-dependent pathogenicity, however, are unknown. In this study, we identify potential virulence mechanisms using whole genome sequencing of V. coralliilyticus ATCC (American Type Culture Collection) BAA-450. Furthermore, we demonstrate direct temperature regulation of numerous virulence factors using proteomic analysis and bioassays. Virulence factors involved in motility, host degradation, secretion, antimicrobial resistance and transcriptional regulation are upregulated at the higher virulent temperature of 27 °C, concurrent with phenotypic changes in motility, antibiotic resistance, hemolysis, cytotoxicity and bioluminescence. These results provide evidence that temperature regulates multiple virulence mechanisms in V. coralliilyticus, independent of abundance. The ecological and biological significance of this temperature-dependent virulence response is reinforced by climate change models that predict tropical SST to consistently exceed 27 °C during the spring, summer and fall seasons. We propose V. coralliilyticus as a model Gram-negative bacterium to study temperature-dependent pathogenicity in Vibrio-related diseases.
Figures
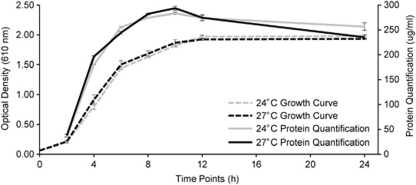
 ) and 27 °C (- - - ). Protein production was measured using the Bradford assay for both 24 °C (
) and 27 °C (- - - ). Protein production was measured using the Bradford assay for both 24 °C (  ) and 27 °C (—) from time points 2–24 h. Error bars represent the standard deviation of three replicate samples.
) and 27 °C (—) from time points 2–24 h. Error bars represent the standard deviation of three replicate samples.

References
-
- Alves N, Neto OSM, Silva BSO, de Moura RL, Francini-Filho RB, Castro CB, et al. Diversity and pathogenic potential of vibrios isolated from Abrolhos Bank corals. Environ Microbiol Rep. 2010;2:90–95. - PubMed
-
- Aronson RB, Precht WF, Macintyre IG, Murdoch TJ. Coral bleach-out in Belize. Nature. 2000;405:36. - PubMed
-
- Arotsker L, Siboni N, Ben-Dov E, Kramarsky-Winter E, Loya Y, Kushmaro A. Vibrio sp. as a potentially important member of the black band disease (BBD) consortium in Favia sp. corals. FEMS Microbiol Ecol. 2009;70:515–524. - PubMed
-
- Austin B, Austin D, Sutherland R, Thompson FL, Swings J. Pathogenicity of vibrios to rainbow trout (Oncorhynchus mykiss, Walbaum) and Artemia nauplii. Environ Microbiol. 2005;7:1488–1495. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
