

Massive analysis of rice small RNAs: mechanistic implications of regulated microRNAs and variants for differential target RNA cleavage
- PMID: 22158467
- PMCID: PMC3269859
- DOI: 10.1105/tpc.111.089045
Massive analysis of rice small RNAs: mechanistic implications of regulated microRNAs and variants for differential target RNA cleavage
Abstract
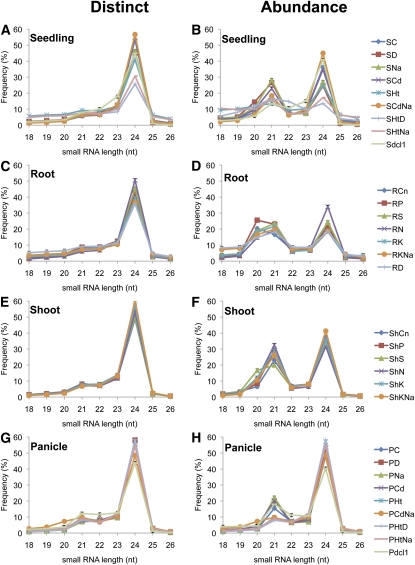
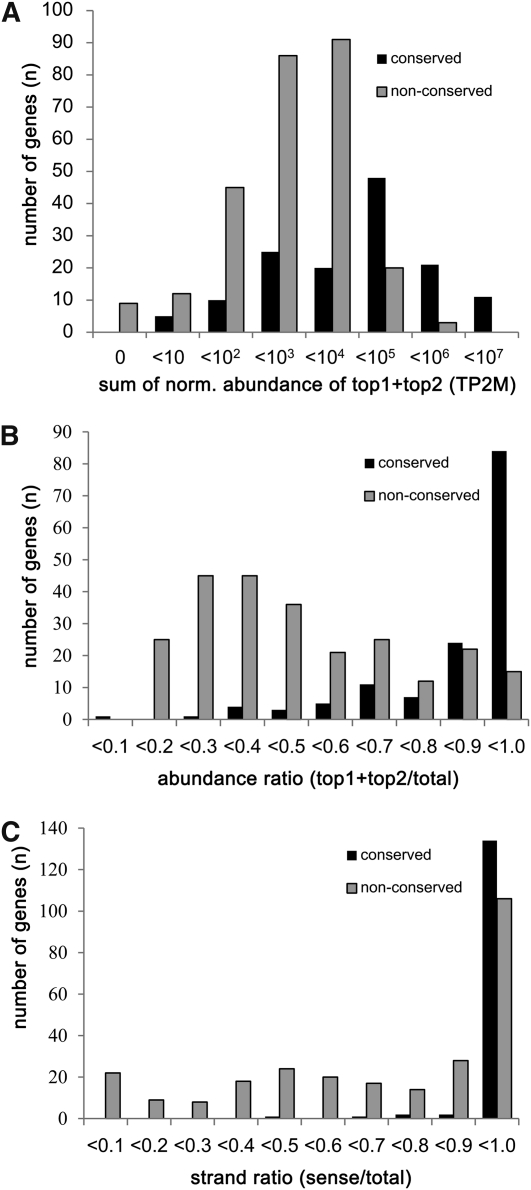
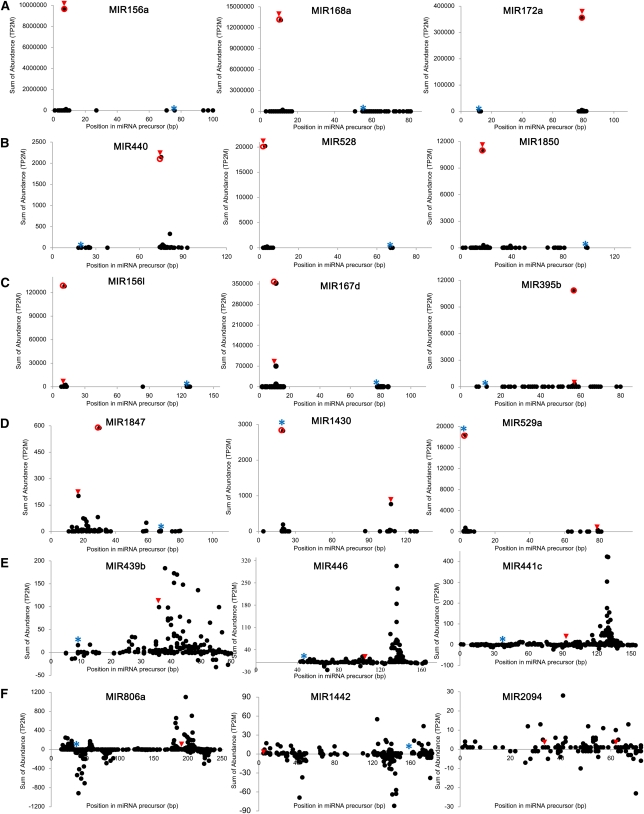
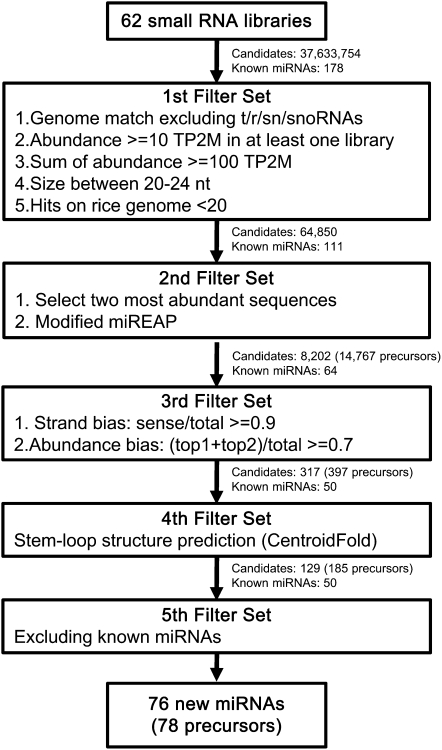
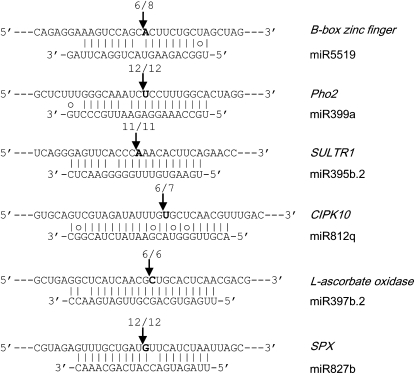
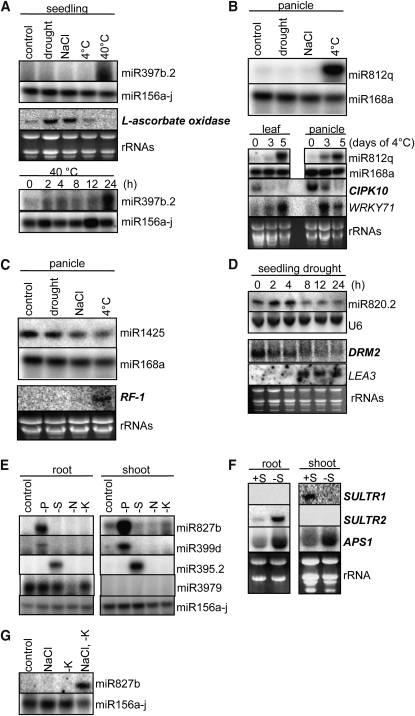
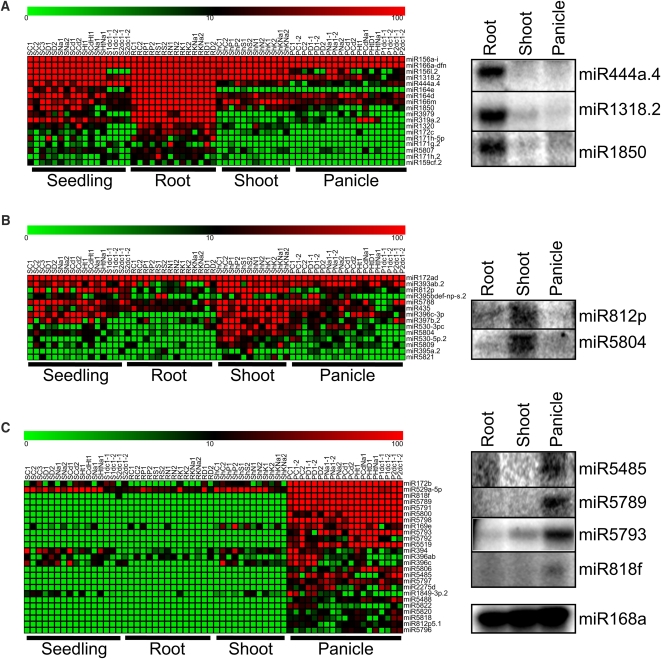
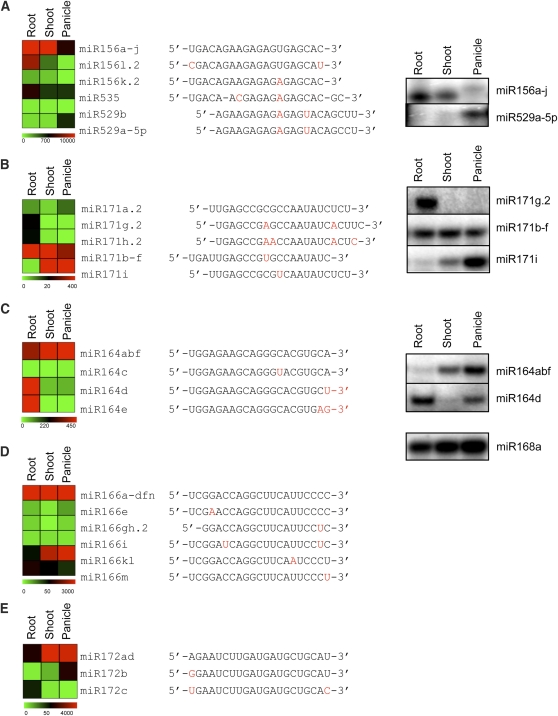
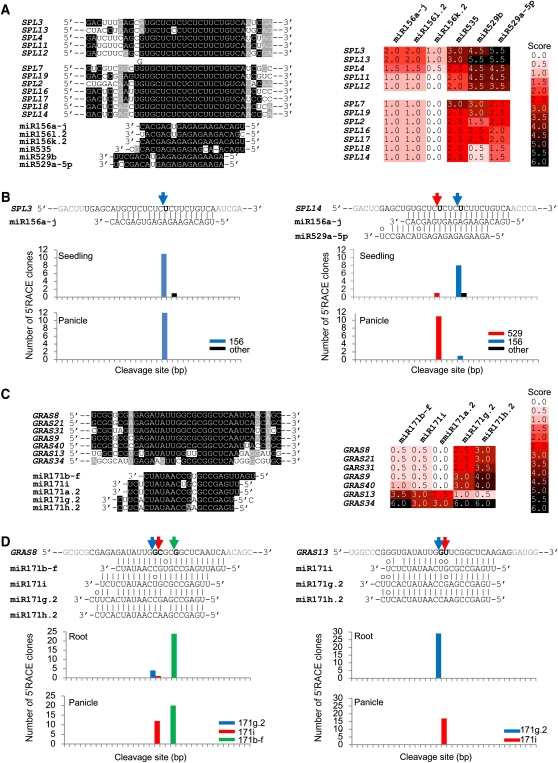
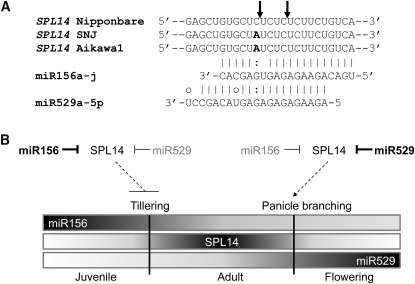
Small RNAs have a variety of important roles in plant development, stress responses, and other processes. They exert their influence by guiding mRNA cleavage, translational repression, and chromatin modification. To identify previously unknown rice (Oryza sativa) microRNAs (miRNAs) and those regulated by environmental stress, 62 small RNA libraries were constructed from rice plants and used for deep sequencing with Illumina technology. The libraries represent several tissues from control plants and plants subjected to different environmental stress treatments. More than 94 million genome-matched reads were obtained, resulting in more than 16 million distinct small RNA sequences. This allowed an evaluation of ~400 annotated miRNAs with current criteria and the finding that among these, ~150 had small interfering RNA-like characteristics. Seventy-six new miRNAs were found, and miRNAs regulated in response to water stress, nutrient stress, or temperature stress were identified. Among the new examples of miRNA regulation were members of the same miRNA family that were differentially regulated in different organs and had distinct sequences Some of these distinct family members result in differential target cleavage and provide new insight about how an agriculturally important rice phenotype could be regulated in the panicle. This high-resolution analysis of rice miRNAs should be relevant to plant miRNAs in general, particularly in the Poaceae.
Figures
References
-
- Arazi T., Talmor-Neiman M., Stav R., Riese M., Huijser P., Baulcombe D.C. (2005). Cloning and characterization of micro-RNAs from moss. Plant J. 43: 837–848 - PubMed
-
- Batistic O., Kudla J. (2009). Plant calcineurin B-like proteins and their interacting protein kinases. Biochim. Biophys. Acta 1793: 985–992 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
