

TCERG1 regulates alternative splicing of the Bcl-x gene by modulating the rate of RNA polymerase II transcription
- PMID: 22158966
- PMCID: PMC3272968
- DOI: 10.1128/MCB.06255-11
TCERG1 regulates alternative splicing of the Bcl-x gene by modulating the rate of RNA polymerase II transcription
Abstract
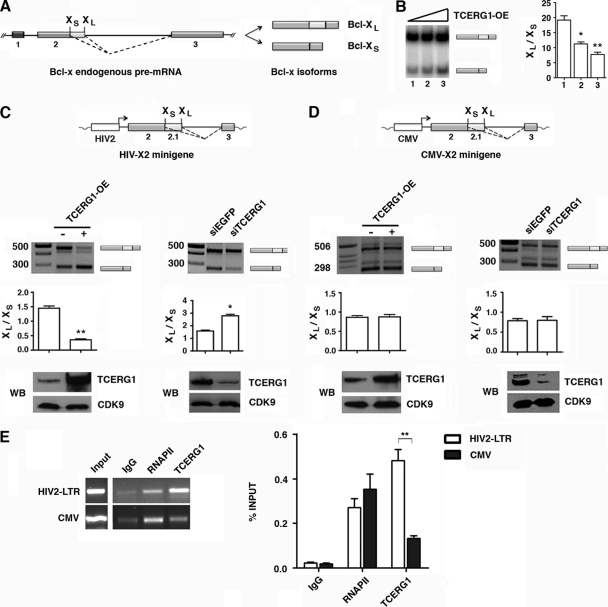
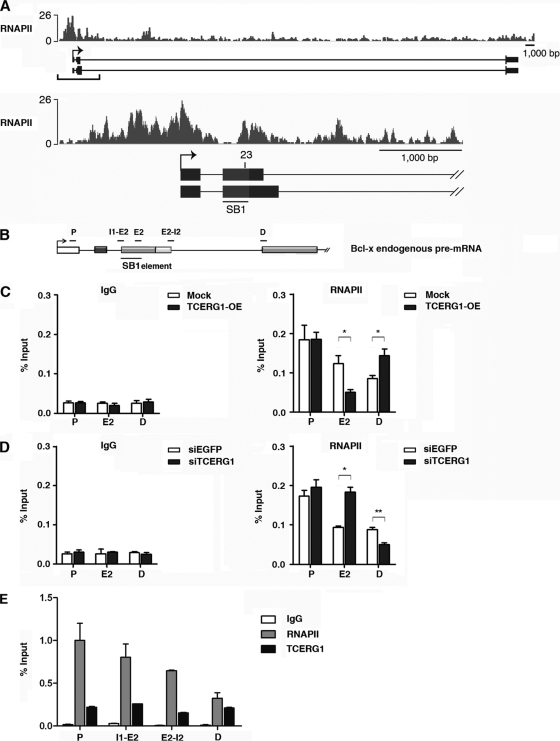
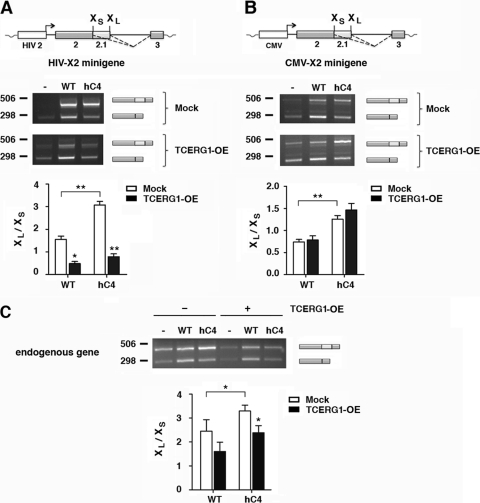
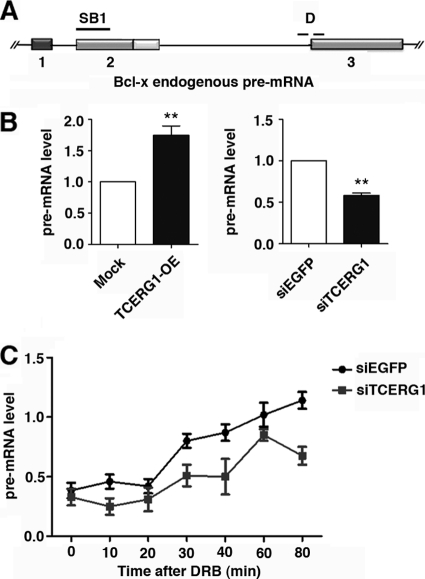
Complex functional coupling exists between transcriptional elongation and pre-mRNA alternative splicing. Pausing sites and changes in the rate of transcription by RNA polymerase II (RNAPII) may therefore have fundamental impacts in the regulation of alternative splicing. Here, we show that the elongation and splicing-related factor TCERG1 regulates alternative splicing of the apoptosis gene Bcl-x in a promoter-dependent manner. TCERG1 promotes the splicing of the short isoform of Bcl-x (Bcl-x(s)) through the SB1 regulatory element located in the first half of exon 2. Consistent with these results, we show that TCERG1 associates with the Bcl-x pre-mRNA. A transcription profile analysis revealed that the RNA sequences required for the effect of TCERG1 on Bcl-x alternative splicing coincide with a putative polymerase pause site. Furthermore, TCERG1 modifies the impact of a slow polymerase on Bcl-x alternative splicing. In support of a role for an elongation mechanism in the transcriptional control of Bcl-x alternative splicing, we found that TCERG1 modifies the amount of pre-mRNAs generated at distal regions of the endogenous Bcl-x. Most importantly, TCERG1 affects the rate of RNAPII transcription of endogenous human Bcl-x. We propose that TCERG1 modulates the elongation rate of RNAPII to relieve pausing, thereby activating the proapoptotic Bcl-x(S) 5' splice site.
Figures

Similar articles
-
Functional Consequences for Apoptosis by Transcription Elongation Regulator 1 (TCERG1)-Mediated Bcl-x and Fas/CD95 Alternative Splicing.PLoS One. 2015 Oct 13;10(10):e0139812. doi: 10.1371/journal.pone.0139812. eCollection 2015. PLoS One. 2015. PMID: 26462236 Free PMC article.
-
Transcription elongation regulator 1 (TCERG1) regulates competent RNA polymerase II-mediated elongation of HIV-1 transcription and facilitates efficient viral replication.Retrovirology. 2013 Oct 28;10:124. doi: 10.1186/1742-4690-10-124. Retrovirology. 2013. PMID: 24165037 Free PMC article.
-
Differential effects of sumoylation on transcription and alternative splicing by transcription elongation regulator 1 (TCERG1).J Biol Chem. 2010 May 14;285(20):15220-15233. doi: 10.1074/jbc.M109.063750. Epub 2010 Mar 9. J Biol Chem. 2010. PMID: 20215116 Free PMC article.
-
Adventures in time and space: splicing efficiency and RNA polymerase II elongation rate.RNA Biol. 2014;11(4):313-9. doi: 10.4161/rna.28646. Epub 2014 Apr 2. RNA Biol. 2014. PMID: 24717535 Free PMC article. Review.
-
Coupling transcription and alternative splicing.Adv Exp Med Biol. 2007;623:175-89. doi: 10.1007/978-0-387-77374-2_11. Adv Exp Med Biol. 2007. PMID: 18380347 Review.
Cited by
-
The chromatin remodeling and mRNA splicing functions of the Brahma (SWI/SNF) complex are mediated by the SNR1/SNF5 regulatory subunit.Nucleic Acids Res. 2012 Jul;40(13):5975-87. doi: 10.1093/nar/gks288. Epub 2012 Mar 29. Nucleic Acids Res. 2012. PMID: 22467207 Free PMC article.
-
MZ1 co-operates with trastuzumab in HER2 positive breast cancer.J Exp Clin Cancer Res. 2021 Mar 19;40(1):106. doi: 10.1186/s13046-021-01907-9. J Exp Clin Cancer Res. 2021. PMID: 33741018 Free PMC article.
-
Aberrant Bcl-x splicing in cancer: from molecular mechanism to therapeutic modulation.J Exp Clin Cancer Res. 2021 Jun 12;40(1):194. doi: 10.1186/s13046-021-02001-w. J Exp Clin Cancer Res. 2021. PMID: 34118966 Free PMC article. Review.
-
Long non-coding RNA INXS is a critical mediator of BCL-XS induced apoptosis.Nucleic Acids Res. 2014 Jul;42(13):8343-55. doi: 10.1093/nar/gku561. Epub 2014 Jul 3. Nucleic Acids Res. 2014. Retraction in: Nucleic Acids Res. 2016 Nov 2;44(19):9518. doi: 10.1093/nar/gkw713. PMID: 24992962 Free PMC article. Retracted.
-
Alternative RNA splicing and cancer.Wiley Interdiscip Rev RNA. 2013 Sep-Oct;4(5):547-66. doi: 10.1002/wrna.1178. Epub 2013 Jun 13. Wiley Interdiscip Rev RNA. 2013. PMID: 23765697 Free PMC article. Review.
References
-
- Allo M, et al. 2009. Control of alternative splicing through siRNA-mediated transcriptional gene silencing. Nat. Struct. Mol. Biol. 16:717–724 - PubMed
-
- Boise LH, et al. 1993. bcl-x, a bcl-2-related gene that functions as a dominant regulator of apoptotic cell death. Cell 74:597–608 - PubMed
-
- Caceres JF, Kornblihtt AR. 2002. Alternative splicing: multiple control mechanisms and involvement in human disease. Trends Genet. 18:186–193 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials