

Cholinergic control in developing prefrontal-hippocampal networks
- PMID: 22159110
- PMCID: PMC6634161
- DOI: 10.1523/JNEUROSCI.2644-11.2011
Cholinergic control in developing prefrontal-hippocampal networks
Abstract
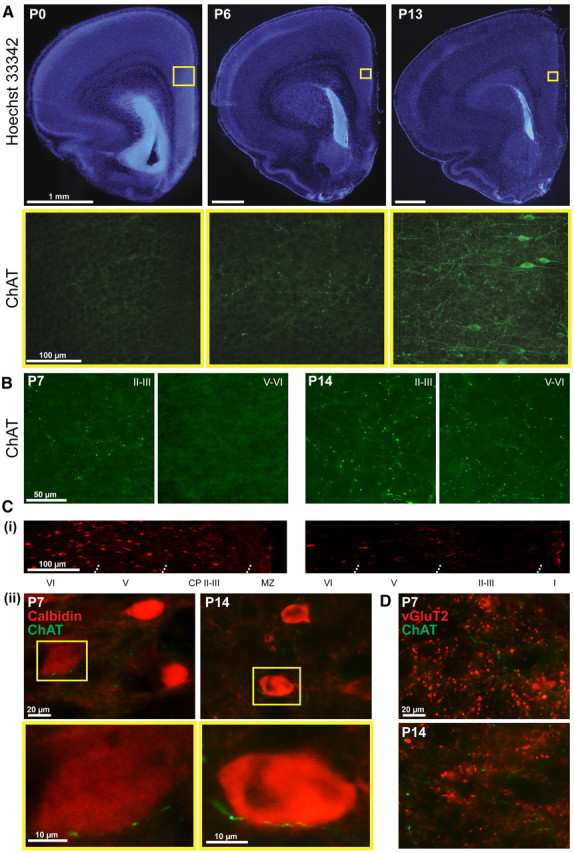
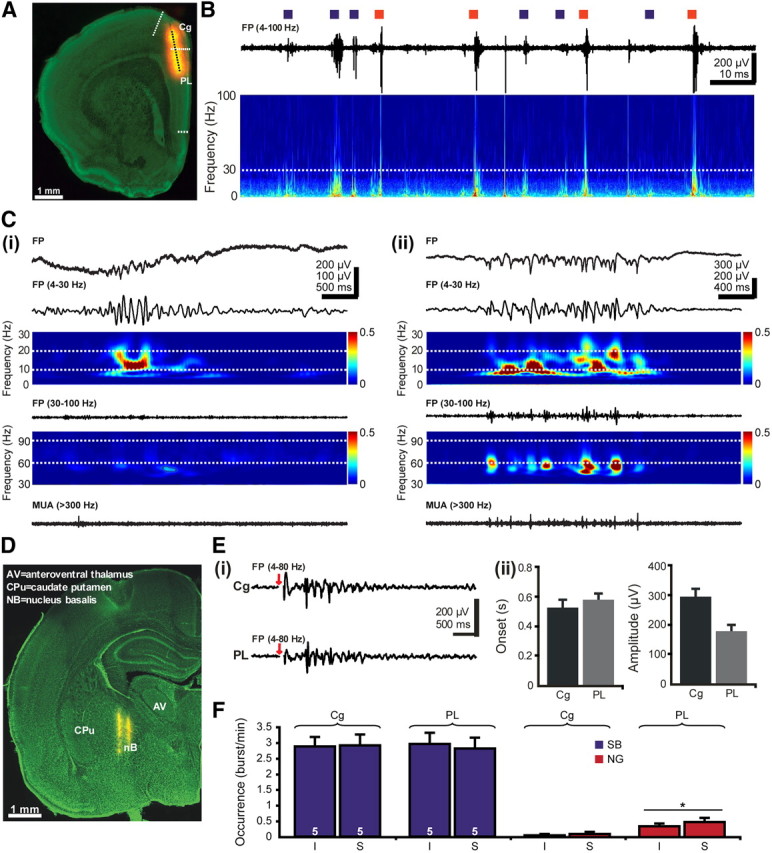
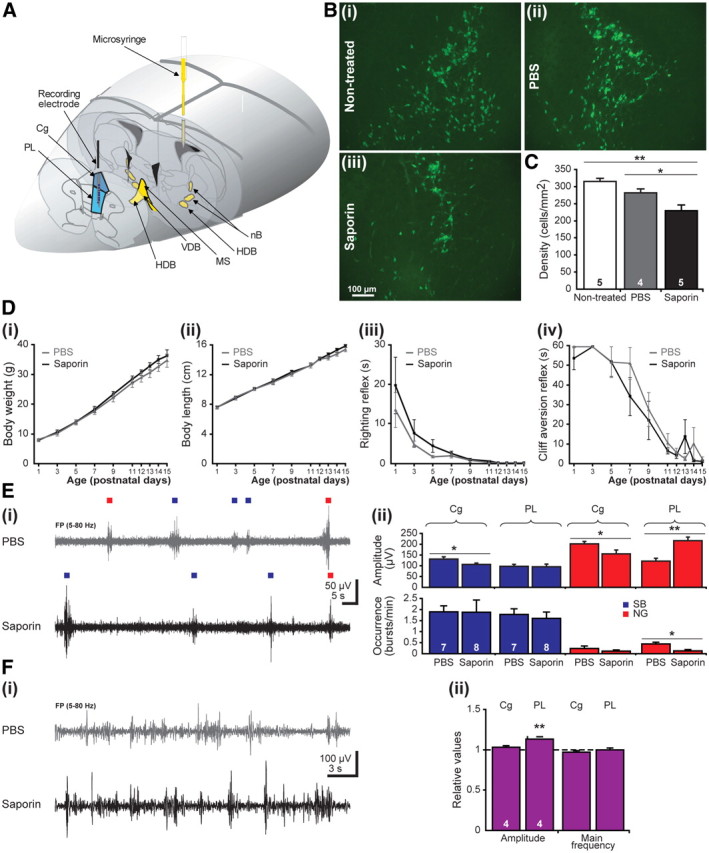
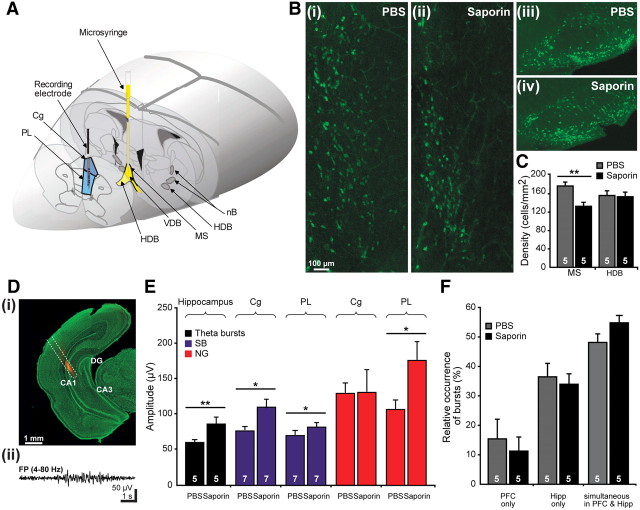
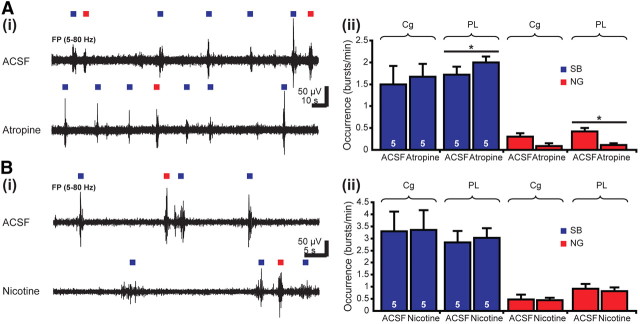
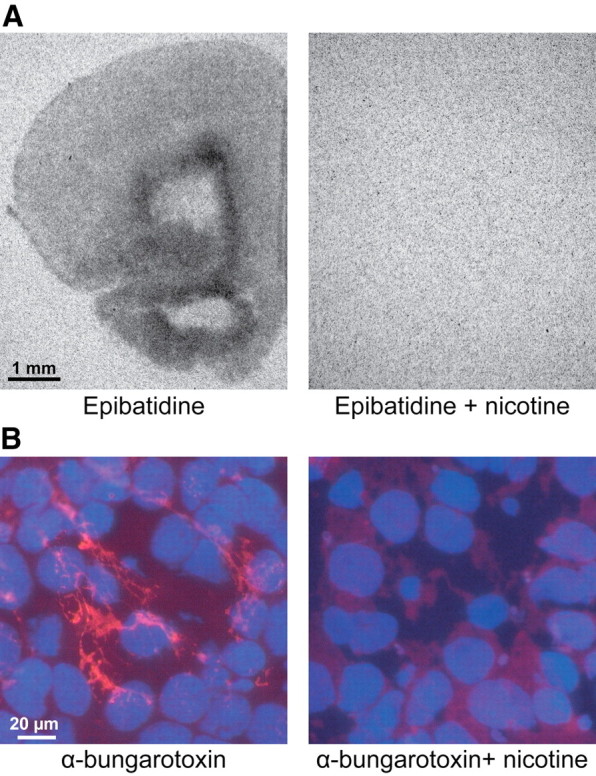
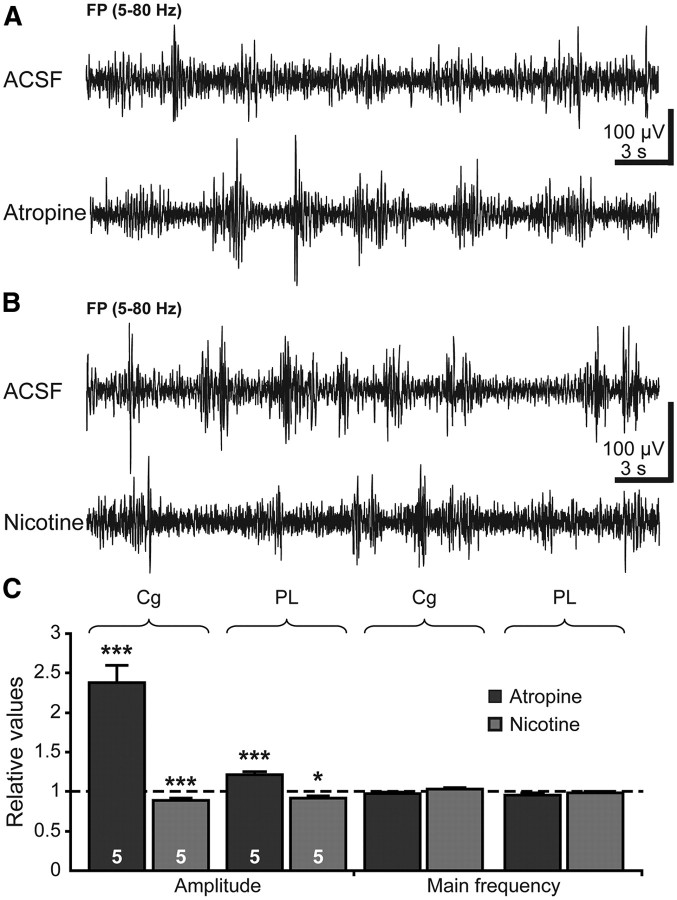
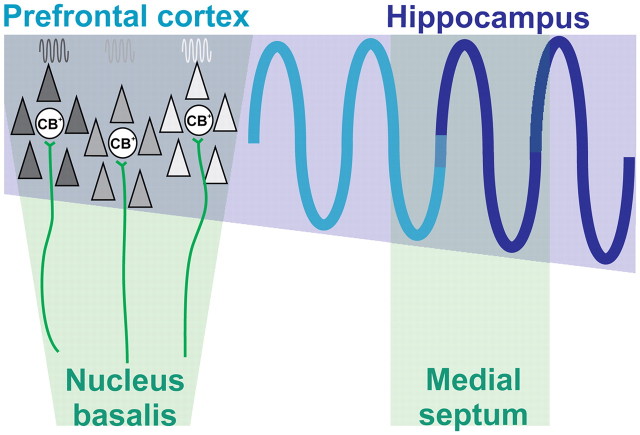
The cholinergic drive enhances input processing in attentional and mnemonic context by interacting with the activity of prefrontal-hippocampal networks. During development, acetylcholine modulates neuronal proliferation, differentiation, and synaptic plasticity, yet its contribution to the maturation of cognitive processing resulting from early entrainment of neuronal networks in oscillatory rhythms remains widely unknown. Here we show that cholinergic projections growing into the rat prefrontal cortex (PFC) toward the end of the first postnatal week boost the generation of nested gamma oscillations superimposed on discontinuous spindle bursts by acting on functional muscarinic but not nicotinic receptors. Although electrical stimulation of cholinergic nuclei increased the occurrence of nested gamma spindle bursts by 41%, diminishment of the cholinergic input by either blockade of the receptors or chronic immunotoxic lesion had the opposite effect. This activation of locally generated gamma episodes by direct cholinergic projections to the PFC was accompanied by indirect modulation of underlying spindle bursts via cholinergic control of hippocampal theta activity. With ongoing maturation and switch of network activity from discontinuous bursts to continuous theta-gamma rhythms, accumulating cholinergic projections acting on both muscarinic and nicotinic receptors mediated the transition from high-amplitude slow to low-amplitude fast rhythms in the PFC. By exerting multiple actions on the oscillatory entrainment of developing prefrontal-hippocampal networks, the cholinergic input may refine them for later gating processing in executive and mnemonic tasks.
Figures
Similar articles
-
Coupled oscillations mediate directed interactions between prefrontal cortex and hippocampus of the neonatal rat.Neuron. 2011 Jul 28;71(2):332-47. doi: 10.1016/j.neuron.2011.05.041. Neuron. 2011. PMID: 21791291
-
Thalamic and Entorhinal Network Activity Differently Modulates the Functional Development of Prefrontal-Hippocampal Interactions.J Neurosci. 2016 Mar 30;36(13):3676-90. doi: 10.1523/JNEUROSCI.3232-15.2016. J Neurosci. 2016. PMID: 27030754 Free PMC article.
-
Cholinergic excitation of septohippocampal GABA but not cholinergic neurons: implications for learning and memory.J Neurosci. 2000 May 15;20(10):3900-8. doi: 10.1523/JNEUROSCI.20-10-03900.2000. J Neurosci. 2000. PMID: 10804229 Free PMC article.
-
Immunolesion by 192IgG-saporin of rat basal forebrain cholinergic system: a useful tool to produce cortical cholinergic dysfunction.Prog Brain Res. 1996;109:253-64. doi: 10.1016/s0079-6123(08)62109-3. Prog Brain Res. 1996. PMID: 9009714 Review.
-
Immunolesion of the cholinergic basal forebrain: effects on functional properties of hippocampal and septal neurons.Int J Dev Neurosci. 1998 Nov-Dec;16(7-8):613-32. doi: 10.1016/s0736-5748(98)00073-2. Int J Dev Neurosci. 1998. PMID: 10198811 Review.
Cited by
-
Knock-Down of Hippocampal DISC1 in Immune-Challenged Mice Impairs the Prefrontal-Hippocampal Coupling and the Cognitive Performance Throughout Development.Cereb Cortex. 2021 Jan 5;31(2):1240-1258. doi: 10.1093/cercor/bhaa291. Cereb Cortex. 2021. PMID: 33037815 Free PMC article.
-
Cholinergic systems are essential for late-stage maturation and refinement of motor cortical circuits.J Neurophysiol. 2015 Mar 1;113(5):1585-97. doi: 10.1152/jn.00408.2014. Epub 2014 Dec 10. J Neurophysiol. 2015. PMID: 25505106 Free PMC article.
-
Developmental dysfunction of prefrontal-hippocampal networks in mouse models of mental illness.Eur J Neurosci. 2019 Sep;50(6):3072-3084. doi: 10.1111/ejn.14436. Epub 2019 Jun 11. Eur J Neurosci. 2019. PMID: 31087437 Free PMC article.
-
Adenosine A(1) receptors in mouse pontine reticular formation depress breathing, increase anesthesia recovery time, and decrease acetylcholine release.Anesthesiology. 2013 Feb;118(2):327-36. doi: 10.1097/ALN.0b013e31827d413e. Anesthesiology. 2013. PMID: 23263018 Free PMC article.
-
Cortical parvalbumin and somatostatin GABA neurons express distinct endogenous modulators of nicotinic acetylcholine receptors.Mol Brain. 2014 Oct 31;7:75. doi: 10.1186/s13041-014-0075-9. Mol Brain. 2014. PMID: 25359633 Free PMC article.
References
-
- Alleva E, Caprioli A, Laviola G. Litter gender composition affects maternal behavior of the primiparous mouse dam (Mus musculus) J Comp Psychol. 1989;103:83–87. - PubMed
-
- Alonso A, Khateb A, Fort P, Jones BE, Mühlethaler M. Differential oscillatory properties of cholinergic and noncholinergic nucleus basalis neurons in guinea pig brain slice. Eur J Neurosci. 1996;8:169–182. - PubMed
-
- Arters J, Hohmann CF, Mills J, Olaghere O, Berger-Sweeney J. Sexually dimorphic responses to neonatal basal forebrain lesions in mice. I. Behavior and neurochemistry. J Neurobiol. 1998;37:582–594. - PubMed
-
- Ashwell K W, Paxinos G. Atlas of the developing rat nervous system. Ed 3. San Diego: Elsevier Academic; 2008.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous