

The kinase Sgg modulates temporal development of macrochaetes in Drosophila by phosphorylation of Scute and Pannier
- PMID: 22159580
- PMCID: PMC3243096
- DOI: 10.1242/dev.074260
The kinase Sgg modulates temporal development of macrochaetes in Drosophila by phosphorylation of Scute and Pannier
Abstract
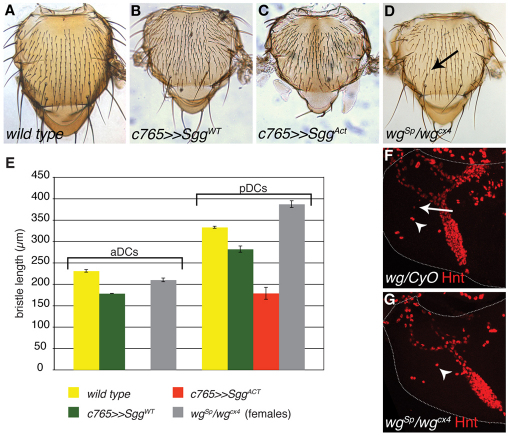
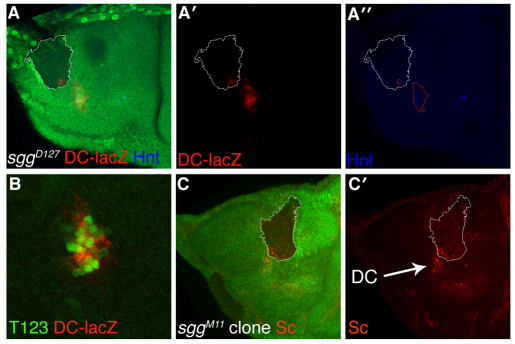
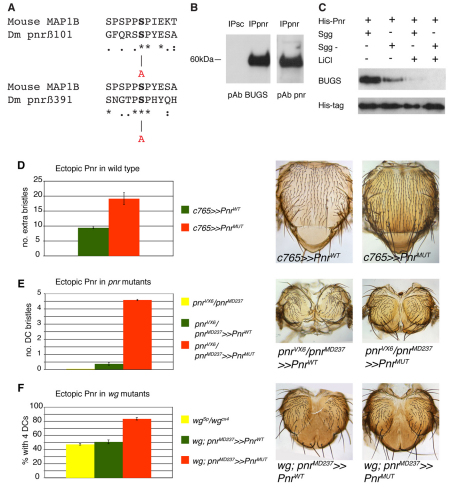
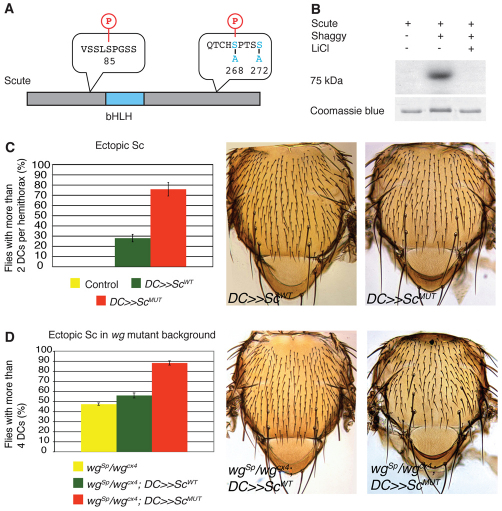
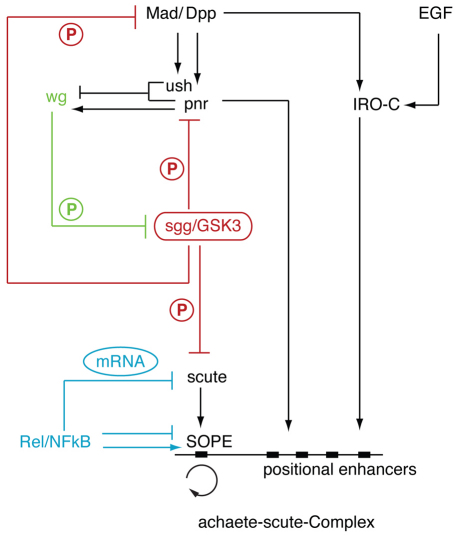
Evolution of novel structures is often made possible by changes in the timing or spatial expression of genes regulating development. Macrochaetes, large sensory bristles arranged into species-specific stereotypical patterns, are an evolutionary novelty of cyclorraphous flies and are associated with changes in both the temporal and spatial expression of the proneural genes achaete (ac) and scute (sc). Changes in spatial expression are associated with the evolution of cis-regulatory sequences, but it is not known how temporal regulation is achieved. One factor required for ac-sc expression, the expression of which coincides temporally with that of ac-sc in the notum, is Wingless (Wg; also known as Wnt). Wingless downregulates the activity of the serine/threonine kinase Shaggy (Sgg; also known as GSK-3). We demonstrate that Scute is phosphorylated by Sgg on a serine residue and that mutation of this residue results in a form of Sc with heightened proneural activity that can rescue the loss of bristles characteristic of wg mutants. We suggest that the phosphorylated form of Sc has reduced transcriptional activity such that sc is unable to autoregulate, an essential function for the segregation of bristle precursors. Sgg also phosphorylates Pannier, a transcriptional activator of ac-sc, the activity of which is similarly dampened when in the phosphorylated state. Furthermore, we show that Wg signalling does not act directly via a cis-regulatory element of the ac-sc genes. We suggest that temporal control of ac-sc activity in cyclorraphous flies is likely to be regulated by permissive factors and might therefore not be encoded at the level of ac-sc gene sequences.
Figures
Similar articles
-
Mutual exclusion of sensory bristles and tendons on the notum of dipteran flies.Curr Biol. 2004 Jun 22;14(12):1047-55. doi: 10.1016/j.cub.2004.06.026. Curr Biol. 2004. PMID: 15202997
-
A conserved trans-regulatory landscape for scute expression on the notum of cyclorraphous Diptera.Dev Genes Evol. 2006 Jan;216(1):29-38. doi: 10.1007/s00427-005-0028-5. Epub 2005 Sep 29. Dev Genes Evol. 2006. PMID: 16193319
-
Drosophila C-terminal binding protein, dCtBP is required for sensory organ prepattern and sharpens proneural transcriptional activity of the GATA factor Pnr.Dev Biol. 2008 Nov 1;323(1):64-75. doi: 10.1016/j.ydbio.2008.08.014. Epub 2008 Aug 22. Dev Biol. 2008. PMID: 18773887
-
Conservation of upstream regulators of scute on the notum of cyclorraphous Diptera.Dev Genes Evol. 2006 Jul-Aug;216(7-8):363-71. doi: 10.1007/s00427-006-0077-4. Epub 2006 May 3. Dev Genes Evol. 2006. PMID: 16670872 Review.
-
How to pattern an epithelium: lessons from achaete-scute regulation on the notum of Drosophila.Gene. 2002 Jun 12;292(1-2):1-12. doi: 10.1016/s0378-1119(02)00628-5. Gene. 2002. PMID: 12119094 Review.
Cited by
-
Apterous A modulates wing size, bristle formation and patterning in Nilaparvata lugens.Sci Rep. 2015 May 21;5:10526. doi: 10.1038/srep10526. Sci Rep. 2015. PMID: 25995006 Free PMC article.
-
Rabex-5 E3 and Rab5 GEF domains differ in their regulation of Ras, Notch, and PI3K signaling in Drosophila wing development.PLoS One. 2024 Oct 28;19(10):e0312274. doi: 10.1371/journal.pone.0312274. eCollection 2024. PLoS One. 2024. PMID: 39466792 Free PMC article.
-
Caspase-dependent non-apoptotic processes in development.Cell Death Differ. 2017 Aug;24(8):1422-1430. doi: 10.1038/cdd.2017.36. Epub 2017 May 19. Cell Death Differ. 2017. PMID: 28524858 Free PMC article. Review.
-
Characterisation of protein isoforms encoded by the Drosophila Glycogen Synthase Kinase 3 gene shaggy.PLoS One. 2020 Aug 6;15(8):e0236679. doi: 10.1371/journal.pone.0236679. eCollection 2020. PLoS One. 2020. PMID: 32760087 Free PMC article.
-
Lysines K117 and K147 play conserved roles in Ras activation from Drosophila to mammals.G3 (Bethesda). 2023 Nov 1;13(11):jkad201. doi: 10.1093/g3journal/jkad201. G3 (Bethesda). 2023. PMID: 37665961 Free PMC article.
References
-
- Berreur P., Porcheron P., Berreur-Bonnenfant J., Simpson P. (1979). Ecdysone levels and pupariation in a temperature sensitive mutation of Drosophila melanogaster. J. Exp. Zool. 210, 347-352
-
- Bertrand N., Castro D. S., Guillemot F. (2002). Proneural genes and the specification of neural cell types. Nat. Rev. Neurosci. 3, 517-530 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
