

Microtubules in bacteria: Ancient tubulins build a five-protofilament homolog of the eukaryotic cytoskeleton
- PMID: 22162949
- PMCID: PMC3232192
- DOI: 10.1371/journal.pbio.1001213
Microtubules in bacteria: Ancient tubulins build a five-protofilament homolog of the eukaryotic cytoskeleton
Abstract
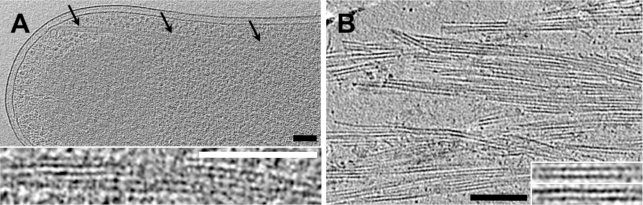
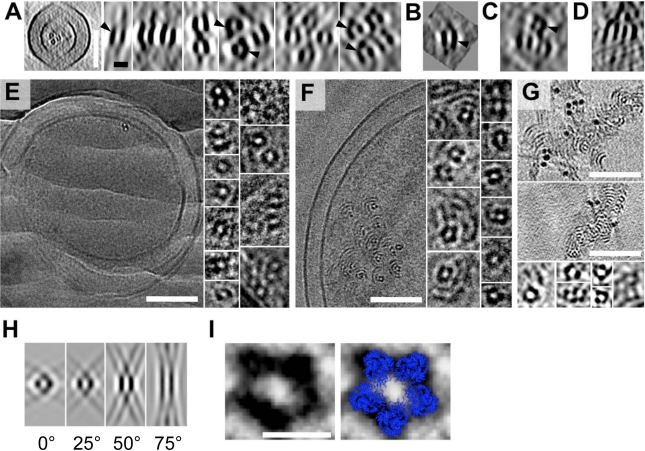
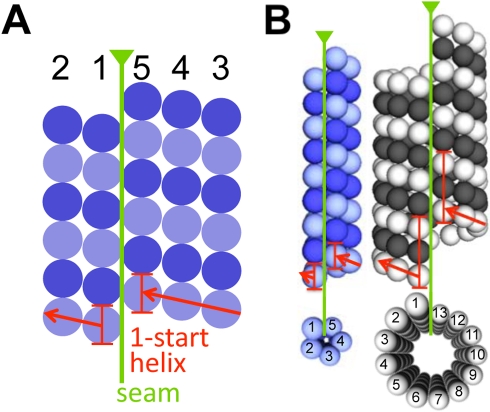
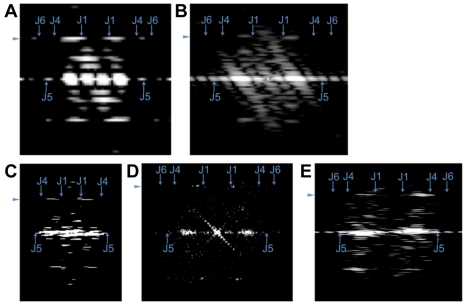
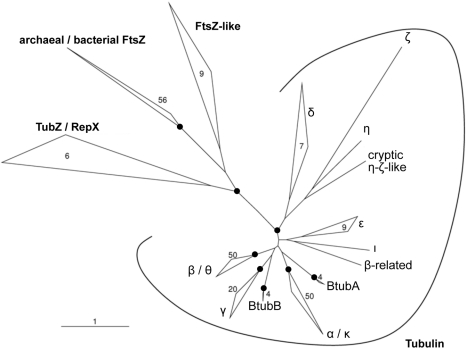
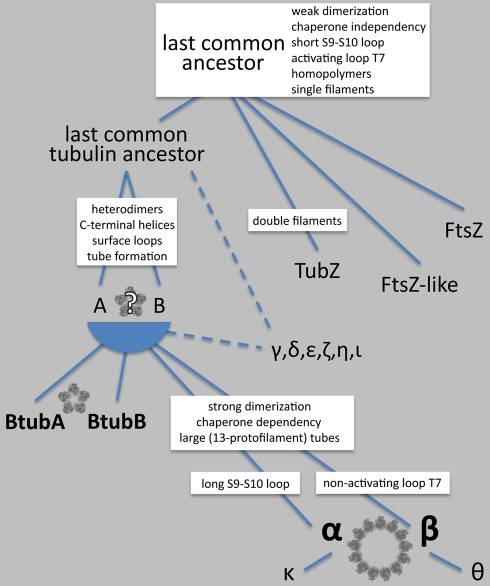
Microtubules play crucial roles in cytokinesis, transport, and motility, and are therefore superb targets for anti-cancer drugs. All tubulins evolved from a common ancestor they share with the distantly related bacterial cell division protein FtsZ, but while eukaryotic tubulins evolved into highly conserved microtubule-forming heterodimers, bacterial FtsZ presumably continued to function as single homopolymeric protofilaments as it does today. Microtubules have not previously been found in bacteria, and we lack insight into their evolution from the tubulin/FtsZ ancestor. Using electron cryomicroscopy, here we show that the tubulin homologs BtubA and BtubB form microtubules in bacteria and suggest these be referred to as "bacterial microtubules" (bMTs). bMTs share important features with their eukaryotic counterparts, such as straight protofilaments and similar protofilament interactions. bMTs are composed of only five protofilaments, however, instead of the 13 typical in eukaryotes. These and other results suggest that rather than being derived from modern eukaryotic tubulin, BtubA and BtubB arose from early tubulin intermediates that formed small microtubules. Since we show that bacterial microtubules can be produced in abundance in vitro without chaperones, they should be useful tools for tubulin research and drug screening.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures

References
-
- Nogales E. Structural insights into microtubule function. Annu Rev Biochem. 2000;69:277–302. - PubMed
-
- Nogales E, Whittaker M, Milligan R. A, Downing K. H. High-resolution model of the microtubule. Cell. 1999;96:79–88. - PubMed
-
- Amos L. A, Schlieper D. Microtubules and maps. Adv Protein Chem. 2005;71:257–298. - PubMed
-
- Erickson H. P. FtsZ, a prokaryotic homolog of tubulin. Cell. 1995;80:367–370. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
