

Sensing-applications of surface-based single vesicle arrays
- PMID: 22163531
- PMCID: PMC3231067
- DOI: 10.3390/s101211352
Sensing-applications of surface-based single vesicle arrays
Abstract
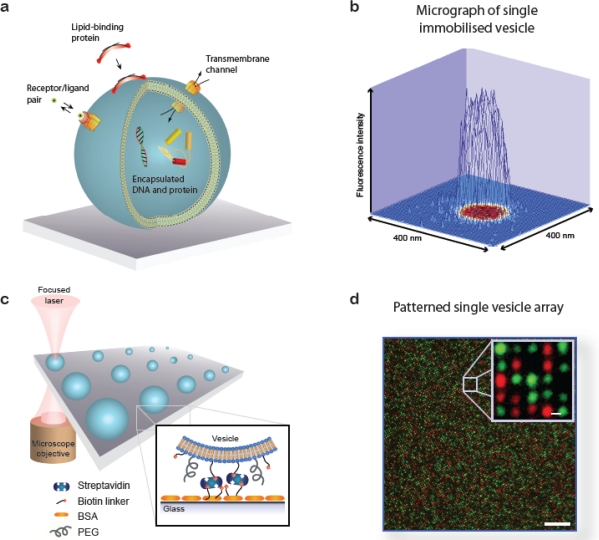
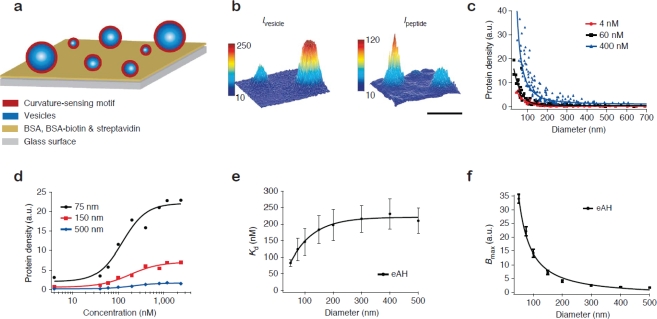
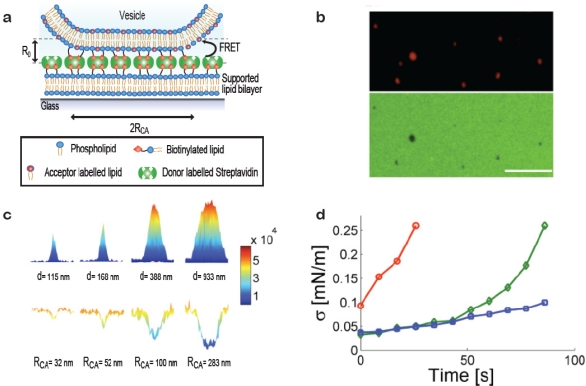
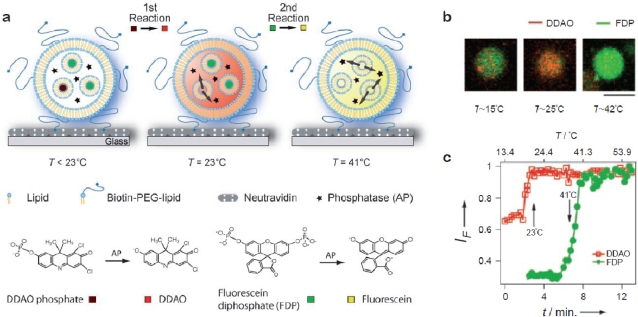
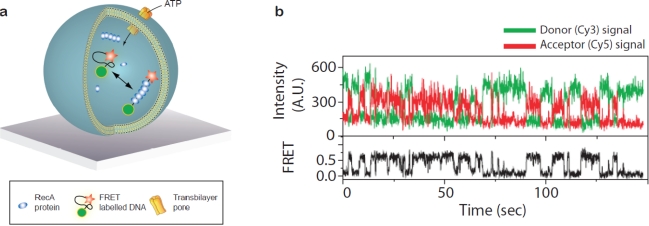
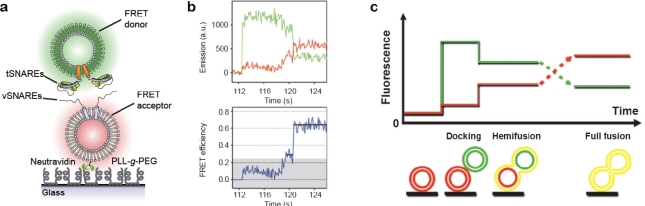
A single lipid vesicle can be regarded as an autonomous ultra-miniaturised 3D biomimetic "scaffold" (Ø≥13 nm) ideally suited for reconstitution and interrogation of biochemical processes. The enclosing lipid bilayer membrane of a vesicle can be applied for studying binding (protein/lipid or receptor/ligand interactions) or transmembrane events (membrane permeability or ion channel activation) while the aqueous vesicle lumen can be used for confining few or single macromolecules and probe, e.g., protein folding, catalytic pathways of enzymes or more complex biochemical reactions, such as signal transduction cascades. Immobilisation (arraying) of single vesicles on a solid support is an extremely useful technique that allows detailed characterisation of vesicle preparations using surface sensitive techniques, in particular fluorescence microscopy. Surface-based single vesicle arrays allow a plethora of prototypic sensing applications in a high throughput format with high spatial and high temporal resolution. In this review we present a series of applications of single vesicle arrays for screening/sensing of: membrane curvature dependent protein-lipid interactions, bilayer tension, reactions triggered in the vesicle lumen, the activity of transmembrane protein channels and biological membrane fusion reactions.
Keywords: model membrane systems; nanoreactors; single vesicles; vesicles.
Figures
References
-
- Stamou D, Duschl C, Delamarche E, Vogel H. Self-Assembled Microarrays of Attoliter Molecular Vessels. Angew Chem Int Ed. 2003;42:5580–5583. - PubMed
-
- Szoka F, Papahadjopoulos D. Comparative Properties and Methods of Preparation of Lipid Vesicles (Liposomes) Annu Rev Biophys Bioeng. 1980;9:467–508. - PubMed
-
- Edwards KA, Baeumner AJ. Analysis of Liposomes. Talanta. 2006;68:1432–1441. - PubMed
-
- Christensen SM, Stamou D. Surface-Based Lipid Vesicle Reactor Systems: Fabrication and Applications. Soft Matter. 2007;3:828–836. - PubMed
-
- Li J, Möhwald H, Ana Z, Lua G. Molecular Assembly of Biomimetic Microcapsules. Soft Matter. 2005;1:259–264. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
