

Neurosteroids and GABA(A) Receptor Interactions: A Focus on Stress
- PMID: 22164129
- PMCID: PMC3230140
- DOI: 10.3389/fnins.2011.00131
Neurosteroids and GABA(A) Receptor Interactions: A Focus on Stress
Abstract
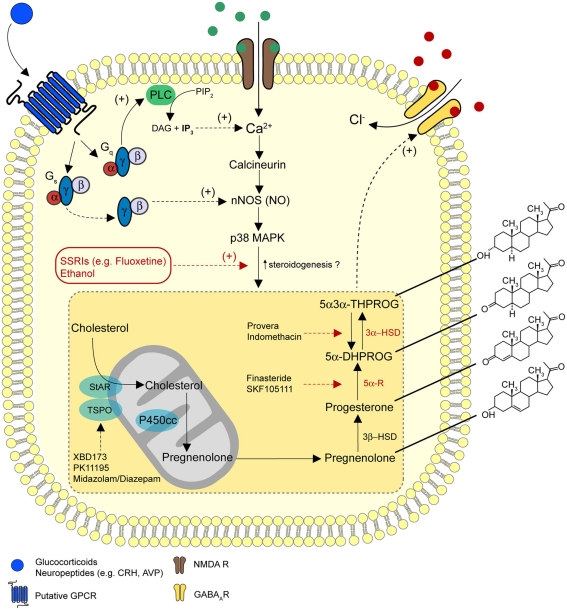
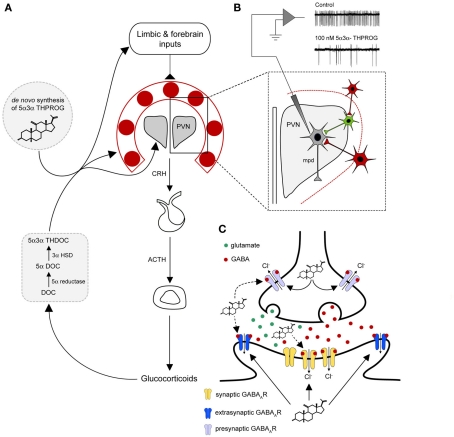
Since the pioneering discovery of the rapid CNS depressant actions of steroids by the "father of stress," Hans Seyle 70 years ago, brain-derived "neurosteroids" have emerged as powerful endogenous modulators of neuronal excitability. The majority of the intervening research has focused on a class of naturally occurring steroids that are metabolites of progesterone and deoxycorticosterone, which act in a non-genomic manner to selectively augment signals mediated by the main inhibitory receptor in the CNS, the GABA(A) receptor. Abnormal levels of such neurosteroids associate with a variety of neurological and psychiatric disorders, suggesting that they serve important physiological and pathophysiological roles. A compelling case can be made to implicate neurosteroids in stress-related disturbances. Here we will critically appraise how brain-derived neurosteroids may impact on the stress response to acute and chronic challenges, both pre- and postnatally through to adulthood. The pathological implications of such actions in the development of psychiatric disturbances will be discussed, with an emphasis on the therapeutic potential of neurosteroids for the treatment of stress-associated disorders.
Keywords: HPA axis; allopregnanolone; anxiety; maternal care; synaptic inhibition.
Figures
References
LinkOut - more resources
Full Text Sources