

Endothelial cell-pericyte interactions stimulate basement membrane matrix assembly: influence on vascular tube remodeling, maturation, and stabilization
- PMID: 22166617
- PMCID: PMC3919655
- DOI: 10.1017/S1431927611012402
Endothelial cell-pericyte interactions stimulate basement membrane matrix assembly: influence on vascular tube remodeling, maturation, and stabilization
Abstract
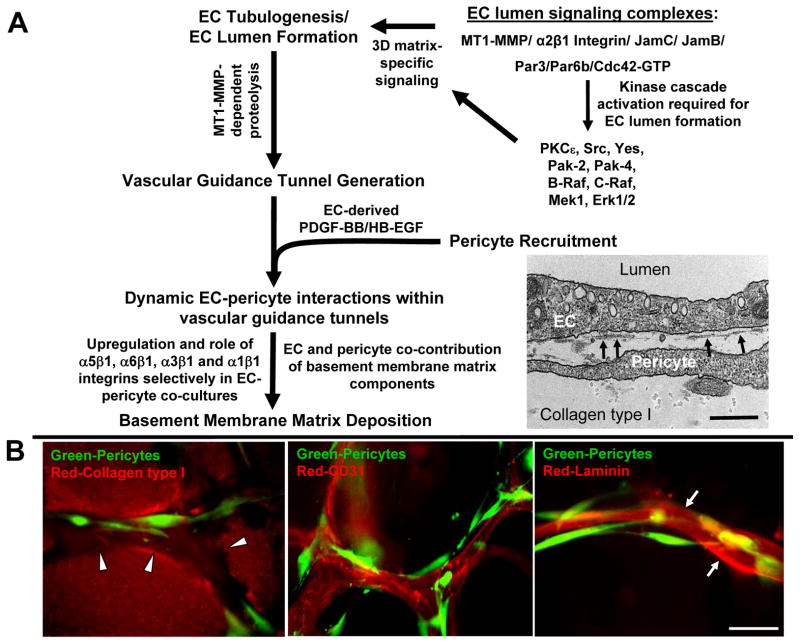
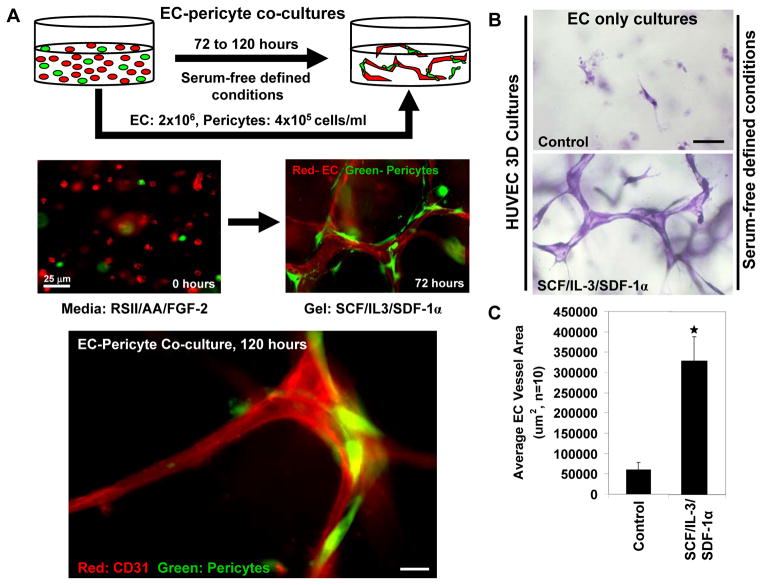
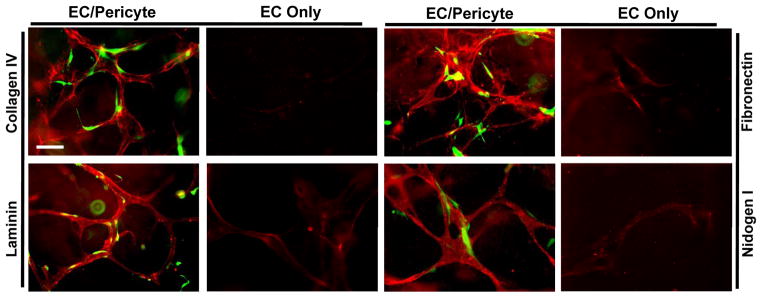
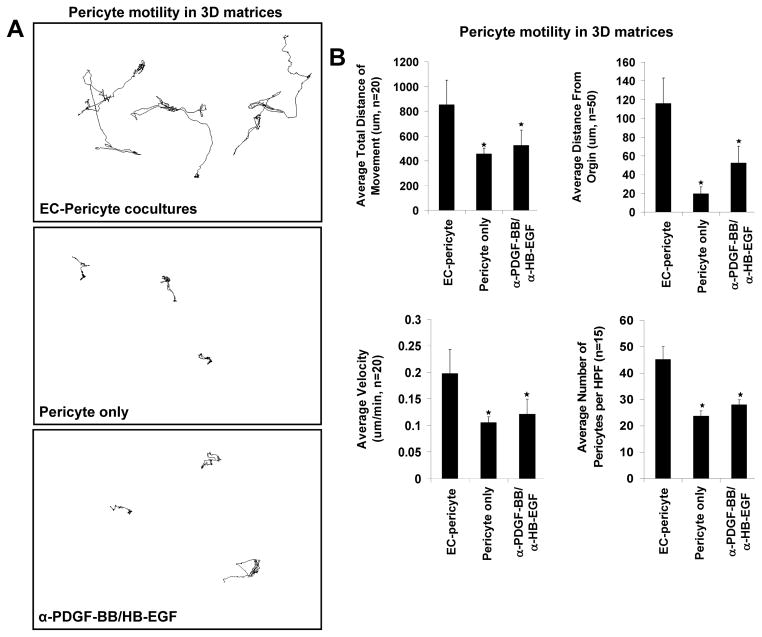
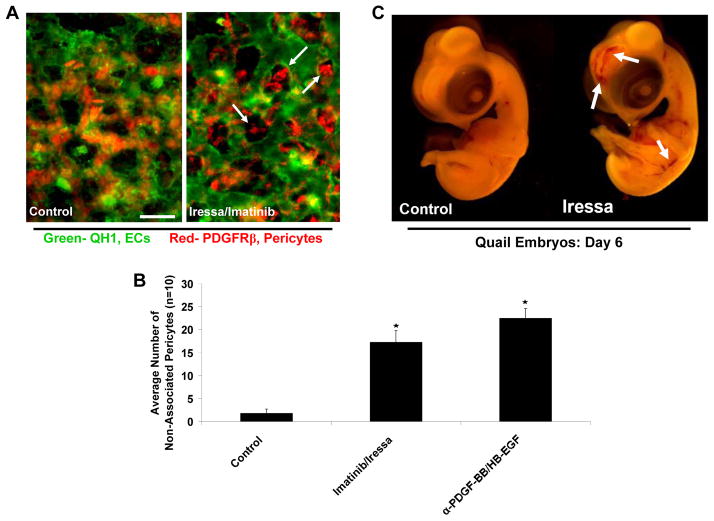
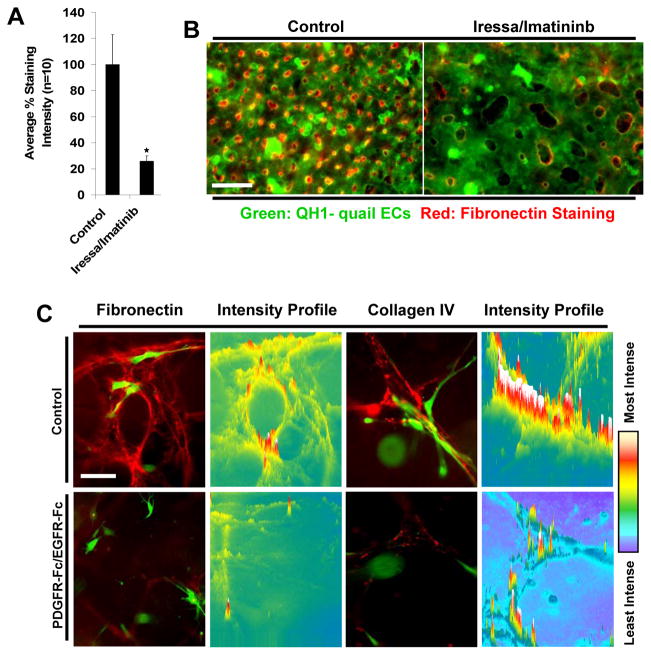
Extracellular matrix synthesis and deposition surrounding the developing vasculature are critical for vessel remodeling and maturation events. Although the basement membrane is an integral structure underlying endothelial cells (ECs), few studies, until recently, have been performed to understand its formation in this context. In this review article, we highlight new data demonstrating a corequirement for ECs and pericytes to properly deposit and assemble vascular basement membranes during morphogenic events. In EC only cultures or under conditions whereby pericyte recruitment is blocked, there is a lack of basement membrane assembly, decreased vessel stability (with increased susceptibility to pro-regressive stimuli), and increased EC tube widths (a marker of dysfunctional EC-pericyte interactions). ECs and pericytes both contribute basement membrane components and, furthermore, both cells induce the expression of particular components as well as integrins that recognize them. The EC-derived factors--platelet derived growth factor-BB and heparin binding-epidermal growth factor--are both critical for pericyte recruitment to EC tubes and concomitant vascular basement membrane formation in vitro and in vivo. Thus, heterotypic EC-pericyte interactions play a fundamental role in vascular basement membrane matrix deposition, a critical tube maturation event that is altered in key disease states such as diabetes and cancer.
Figures
References
-
- Adams RH, Alitalo K. Molecular regulation of angiogenesis and lymphangiogenesis. Nat Rev Mol Cell Biol. 2007;8(6):464–78. - PubMed
