

Catalytic mechanism of Sep-tRNA:Cys-tRNA synthase: sulfur transfer is mediated by disulfide and persulfide
- PMID: 22167197
- PMCID: PMC3285321
- DOI: 10.1074/jbc.M111.313700
Catalytic mechanism of Sep-tRNA:Cys-tRNA synthase: sulfur transfer is mediated by disulfide and persulfide
Abstract
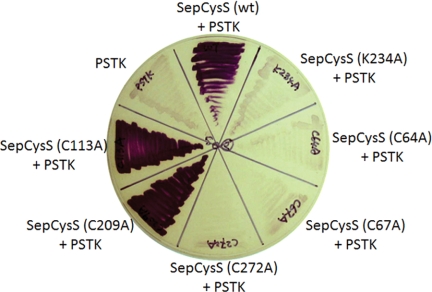
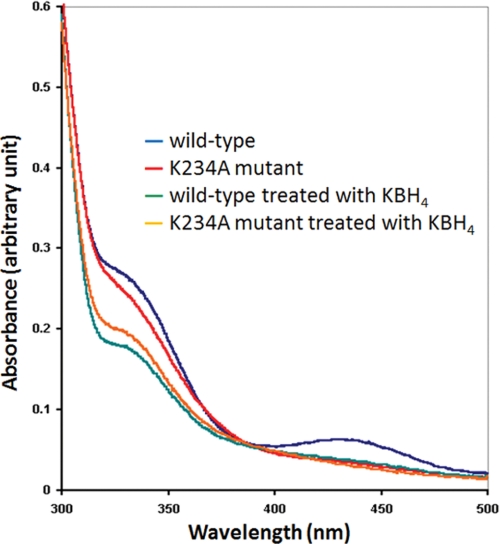
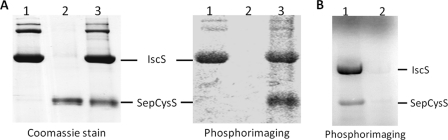
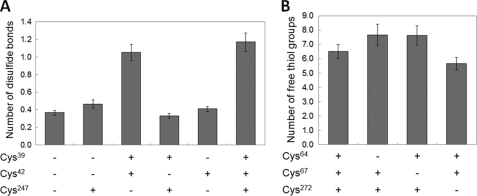
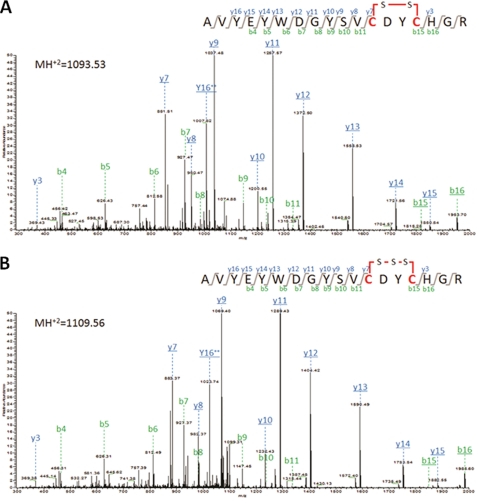
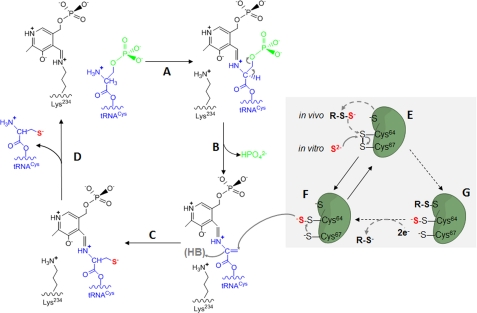
Sep-tRNA:Cys-tRNA synthase (SepCysS) catalyzes the sulfhydrylation of tRNA-bound O-phosphoserine (Sep) to form cysteinyl-tRNA(Cys) (Cys-tRNA(Cys)) in methanogens that lack the canonical cysteinyl-tRNA synthetase (CysRS). A crystal structure of the Archaeoglobus fulgidus SepCysS apoenzyme provides information on the binding of the pyridoxal phosphate cofactor as well as on amino acid residues that may be involved in substrate binding. However, the mechanism of sulfur transfer to form cysteine was not known. Using an in vivo Escherichia coli complementation assay, we showed that all three highly conserved Cys residues in SepCysS (Cys(64), Cys(67), and Cys(272) in the Methanocaldococcus jannaschii enzyme) are essential for the sulfhydrylation reaction in vivo. Biochemical and mass spectrometric analysis demonstrated that Cys(64) and Cys(67) form a disulfide linkage and carry a sulfane sulfur in a portion of the enzyme. These results suggest that a persulfide group (containing a sulfane sulfur) is the proximal sulfur donor for cysteine biosynthesis. The presence of Cys(272) increased the amount of sulfane sulfur in SepCysS by 3-fold, suggesting that this Cys residue facilitates the generation of the persulfide group. Based upon these findings, we propose for SepCysS a sulfur relay mechanism that recruits both disulfide and persulfide intermediates.
Figures
Similar articles
-
Structural insights into the second step of RNA-dependent cysteine biosynthesis in archaea: crystal structure of Sep-tRNA:Cys-tRNA synthase from Archaeoglobus fulgidus.J Mol Biol. 2007 Jun 29;370(1):128-41. doi: 10.1016/j.jmb.2007.04.050. Epub 2007 May 4. J Mol Biol. 2007. PMID: 17512006
-
RNA-dependent cysteine biosynthesis in archaea.Science. 2005 Mar 25;307(5717):1969-72. doi: 10.1126/science.1108329. Science. 2005. PMID: 15790858
-
RNA-Dependent Cysteine Biosynthesis in Bacteria and Archaea.mBio. 2017 May 9;8(3):e00561-17. doi: 10.1128/mBio.00561-17. mBio. 2017. PMID: 28487430 Free PMC article.
-
Enzymatic Regulation and Biological Functions of Reactive Cysteine Persulfides and Polysulfides.Biomolecules. 2020 Aug 27;10(9):1245. doi: 10.3390/biom10091245. Biomolecules. 2020. PMID: 32867265 Free PMC article. Review.
-
Persulfide synthases that are functionally coupled with translation mediate sulfur respiration in mammalian cells.Br J Pharmacol. 2019 Feb;176(4):607-615. doi: 10.1111/bph.14356. Epub 2018 Jun 7. Br J Pharmacol. 2019. PMID: 29748969 Free PMC article. Review.
Cited by
-
A [3Fe-4S] cluster is required for tRNA thiolation in archaea and eukaryotes.Proc Natl Acad Sci U S A. 2016 Nov 8;113(45):12703-12708. doi: 10.1073/pnas.1615732113. Epub 2016 Oct 24. Proc Natl Acad Sci U S A. 2016. PMID: 27791189 Free PMC article.
-
Ancient translation factor is essential for tRNA-dependent cysteine biosynthesis in methanogenic archaea.Proc Natl Acad Sci U S A. 2014 Jul 22;111(29):10520-5. doi: 10.1073/pnas.1411267111. Epub 2014 Jul 7. Proc Natl Acad Sci U S A. 2014. PMID: 25002468 Free PMC article.
-
The putative tRNA 2-thiouridine synthetase Ncs6 is an essential sulfur carrier in Methanococcus maripaludis.FEBS Lett. 2014 Mar 18;588(6):873-7. doi: 10.1016/j.febslet.2014.01.065. Epub 2014 Feb 11. FEBS Lett. 2014. PMID: 24530533 Free PMC article.
-
Interplay between oxygen and Fe-S cluster biogenesis: insights from the Suf pathway.Biochemistry. 2014 Sep 23;53(37):5834-47. doi: 10.1021/bi500488r. Epub 2014 Sep 11. Biochemistry. 2014. PMID: 25153801 Free PMC article. Review.
-
Efficient Sulfide Assimilation in Methanosarcina acetivorans Is Mediated by the MA1715 Protein.J Bacteriol. 2016 Jun 27;198(14):1974-83. doi: 10.1128/JB.00141-16. Print 2016 Jul 15. J Bacteriol. 2016. PMID: 27137504 Free PMC article.
References
-
- Sauerwald A., Zhu W., Major T. A., Roy H., Palioura S., Jahn D., Whitman W. B., Yates J. R., 3rd, Ibba M., Söll D. (2005) RNA-dependent cysteine biosynthesis in archaea. Science 307, 1969–1972 - PubMed
-
- Li T., Graham D. E., Stathopoulos C., Haney P. J., Kim H. S., Vothknecht U., Kitabatake M., Hong K. W., Eggertsson G., Curnow A. W., Lin W., Celic I., Whitman W., Söll D. (1999) Cysteinyl-tRNA formation: the last puzzle of aminoacyl-tRNA synthesis. FEBS Lett. 462, 302–306 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
