

Calcium-induced folding and stabilization of the Pseudomonas aeruginosa alkaline protease
- PMID: 22170064
- PMCID: PMC3281706
- DOI: 10.1074/jbc.M111.310300
Calcium-induced folding and stabilization of the Pseudomonas aeruginosa alkaline protease
Abstract
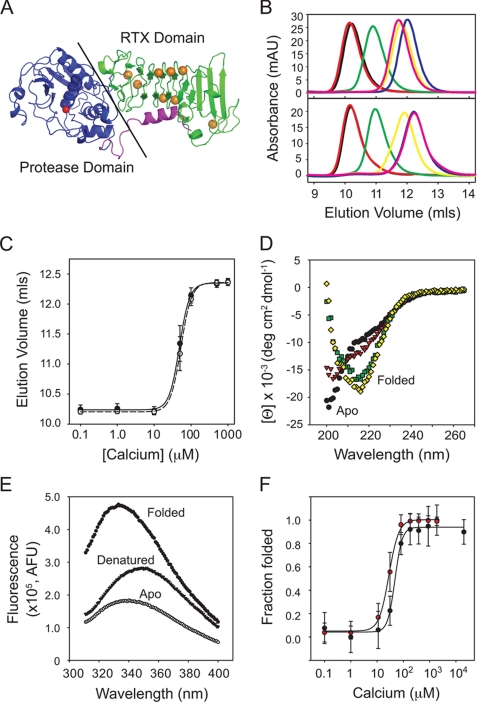
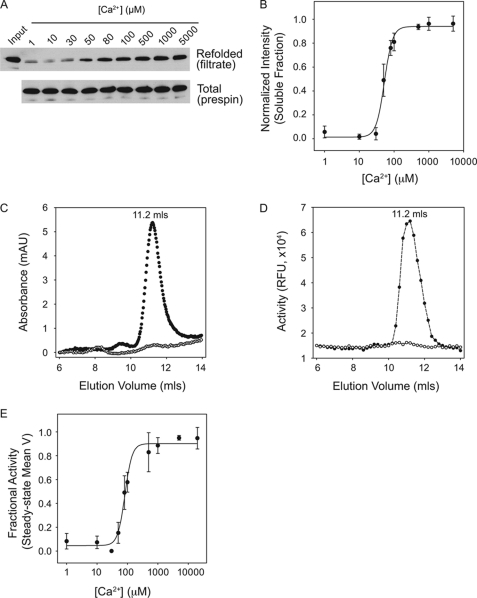
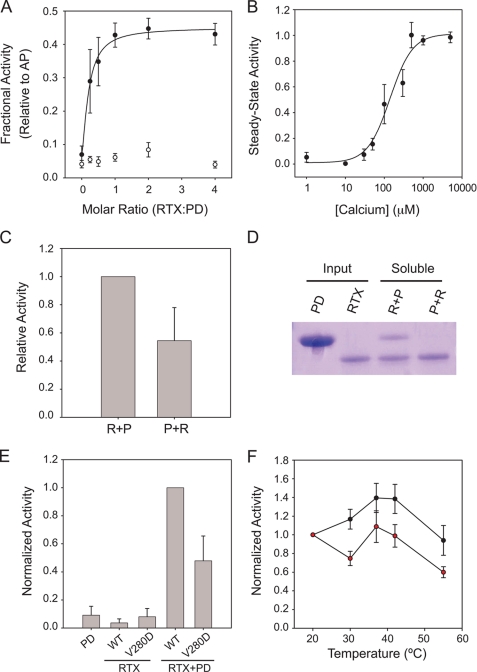
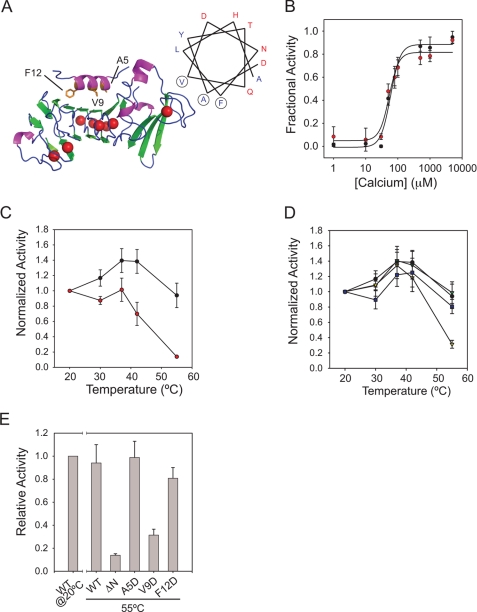
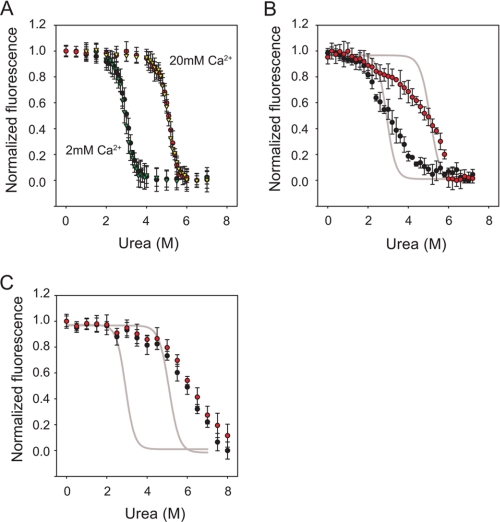
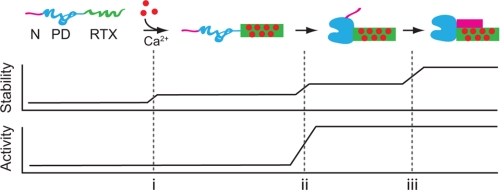
Pseudomonas aeruginosa is an opportunistic pathogen that contributes to the mortality of immunocompromised individuals and patients with cystic fibrosis. Pseudomonas infection presents clinical challenges due to its ability to form biofilms and modulate host-pathogen interactions through the secretion of virulence factors. The calcium-regulated alkaline protease (AP), a member of the repeats in toxin (RTX) family of proteins, is implicated in multiple modes of infection. A series of full-length and truncation mutants were purified for structural and functional studies to evaluate the role of Ca(2+) in AP folding and activation. We find that Ca(2+) binding induces RTX folding, which serves to chaperone the folding of the protease domain. Subsequent association of the RTX domain with an N-terminal α-helix stabilizes AP. These results provide a basis for the Ca(2+)-mediated regulation of AP and suggest mechanisms by which Ca(2+) regulates the RTX family of virulence factors.
Figures
Similar articles
-
Activation of the epithelial sodium channel (ENaC) by the alkaline protease from Pseudomonas aeruginosa.J Biol Chem. 2012 Sep 21;287(39):32556-65. doi: 10.1074/jbc.M112.369520. Epub 2012 Aug 2. J Biol Chem. 2012. PMID: 22859302 Free PMC article.
-
Inhibition of Pseudomonas aeruginosa virulence: characterization of the AprA-AprI interface and species selectivity.J Mol Biol. 2012 Jan 20;415(3):573-83. doi: 10.1016/j.jmb.2011.11.039. Epub 2011 Nov 29. J Mol Biol. 2012. PMID: 22154939
-
The Pseudomonas aeruginosa PAO1 Two-Component Regulator CarSR Regulates Calcium Homeostasis and Calcium-Induced Virulence Factor Production through Its Regulatory Targets CarO and CarP.J Bacteriol. 2016 Jan 11;198(6):951-63. doi: 10.1128/JB.00963-15. J Bacteriol. 2016. PMID: 26755627 Free PMC article.
-
[Molecular determinants in regulating Pseudomonas aeruginosa type III secretion system--a review].Wei Sheng Wu Xue Bao. 2008 Oct;48(10):1413-7. Wei Sheng Wu Xue Bao. 2008. PMID: 19160828 Review. Chinese.
-
Cytoplasmic molecular chaperones in Pseudomonas species.J Microbiol. 2022 Nov;60(11):1049-1060. doi: 10.1007/s12275-022-2425-0. Epub 2022 Nov 1. J Microbiol. 2022. PMID: 36318358 Review.
Cited by
-
A serralysin-like protein of Candidatus Liberibacter asiaticus modulates components of the bacterial extracellular matrix.Front Microbiol. 2022 Oct 19;13:1006962. doi: 10.3389/fmicb.2022.1006962. eCollection 2022. Front Microbiol. 2022. PMID: 36338045 Free PMC article.
-
Inducible polymerization and two-dimensional assembly of the repeats-in-toxin (RTX) domain from the Pseudomonas aeruginosa alkaline protease.Biochemistry. 2014 Oct 21;53(41):6452-62. doi: 10.1021/bi5007546. Epub 2014 Oct 3. Biochemistry. 2014. PMID: 25232897 Free PMC article.
-
Draft genome sequence of Pseudomonas sp. strain M47T1, carried by Bursaphelenchus xylophilus isolated from Pinus pinaster.J Bacteriol. 2012 Sep;194(17):4789-90. doi: 10.1128/JB.01116-12. J Bacteriol. 2012. PMID: 22887683 Free PMC article.
-
Identification of SlpB, a Cytotoxic Protease from Serratia marcescens.Infect Immun. 2015 Jul;83(7):2907-16. doi: 10.1128/IAI.03096-14. Epub 2015 May 4. Infect Immun. 2015. PMID: 25939509 Free PMC article.
-
Receptor-Based Peptides for Inhibition of Leukotoxin Activity.ACS Infect Dis. 2018 Jul 13;4(7):1073-1081. doi: 10.1021/acsinfecdis.7b00230. Epub 2018 May 17. ACS Infect Dis. 2018. PMID: 29742342 Free PMC article.
References
-
- Lyczak J. B., Cannon C. L., Pier G. B. (2000) Establishment of Pseudomonas aeruginosa infection. Lessons from a versatile opportunist. Microbes Infect. 2, 1051–1060 - PubMed
-
- Hobden J. A. (2002) Pseudomonas aeruginosa proteases and corneal virulence. DNA Cell Biol. 21, 391–396 - PubMed
-
- Leidal K. G., Munson K. L., Johnson M. C., Denning G. M. (2003) Metalloproteases from Pseudomonas aeruginosa degrade human RANTES, MCP-1, and ENA-78. J. Interferon Cytokine Res. 23, 307–318 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
