

Nitrite oxidation in the Namibian oxygen minimum zone
- PMID: 22170426
- PMCID: PMC3358024
- DOI: 10.1038/ismej.2011.178
Nitrite oxidation in the Namibian oxygen minimum zone
Abstract
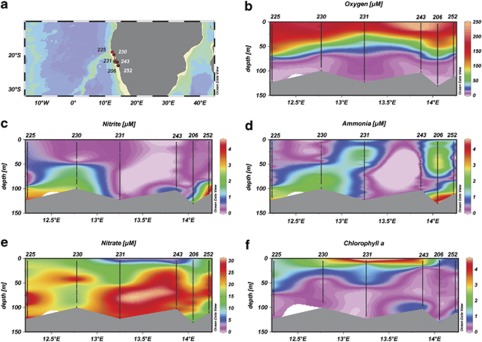
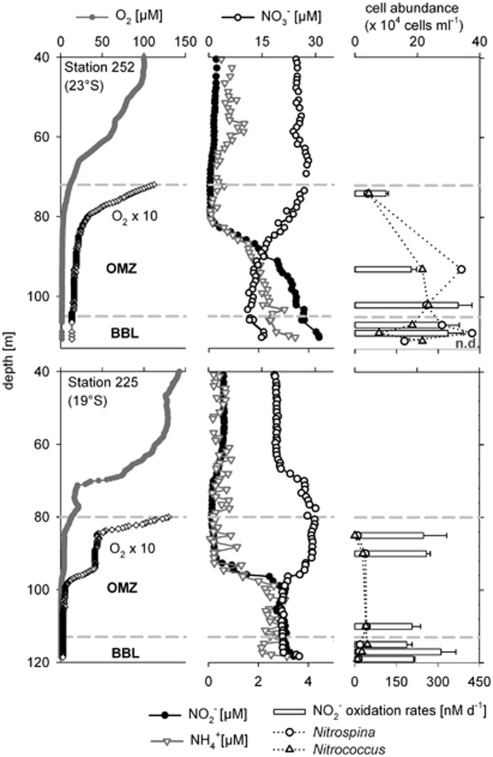
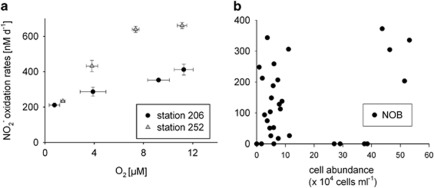
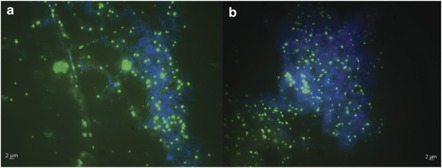
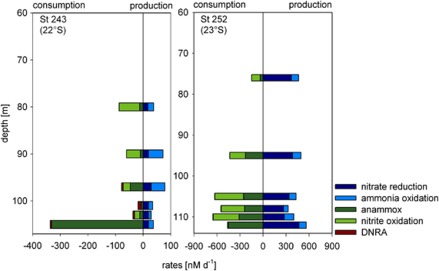
Nitrite oxidation is the second step of nitrification. It is the primary source of oceanic nitrate, the predominant form of bioavailable nitrogen in the ocean. Despite its obvious importance, nitrite oxidation has rarely been investigated in marine settings. We determined nitrite oxidation rates directly in (15)N-incubation experiments and compared the rates with those of nitrate reduction to nitrite, ammonia oxidation, anammox, denitrification, as well as dissimilatory nitrate/nitrite reduction to ammonium in the Namibian oxygen minimum zone (OMZ). Nitrite oxidation (≤372 nM NO(2)(-) d(-1)) was detected throughout the OMZ even when in situ oxygen concentrations were low to non-detectable. Nitrite oxidation rates often exceeded ammonia oxidation rates, whereas nitrate reduction served as an alternative and significant source of nitrite. Nitrite oxidation and anammox co-occurred in these oxygen-deficient waters, suggesting that nitrite-oxidizing bacteria (NOB) likely compete with anammox bacteria for nitrite when substrate availability became low. Among all of the known NOB genera targeted via catalyzed reporter deposition fluorescence in situ hybridization, only Nitrospina and Nitrococcus were detectable in the Namibian OMZ samples investigated. These NOB were abundant throughout the OMZ and contributed up to ~9% of total microbial community. Our combined results reveal that a considerable fraction of the recently recycled nitrogen or reduced NO(3)(-) was re-oxidized back to NO(3)(-) via nitrite oxidation, instead of being lost from the system through the anammox or denitrification pathways.
Figures
References
-
- Alawi M, Lipski A, Sanders T, Eva Maria P, Spieck E. Cultivation of a novel cold-adapted nitrite oxidizing betaproteobacterium from the Siberian Arctic. ISME J. 2007;1:256–264. - PubMed
-
- Beman JM, Sachdeva R, Fuhrman JA. Population ecology of nitrifying archaea and bacteria in the southern California bight. Environ Microbiol. 2010;12:1282–1292. - PubMed
-
- Codispoti LA, Yoshinari T, Devol AH.2005Suboxic respiration in the oceanic water columnIn: Giorgio P del, Williams PJ LeB (eds). Respiration in Aquatic Ecosystems Oxford University Press: Oxford; 225–247.
-
- Dalsgaard T, Canfield DE, Petersen J, Thamdrup B, Acuna-Gonzalez J. N-2 production by the anammox reaction in the anoxic water column of Golfo Dulce, Costa Rica. Nature. 2003;422:606–608. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
