

Perforin activity at membranes leads to invaginations and vesicle formation
- PMID: 22173634
- PMCID: PMC3248536
- DOI: 10.1073/pnas.1107473108
Perforin activity at membranes leads to invaginations and vesicle formation
Abstract
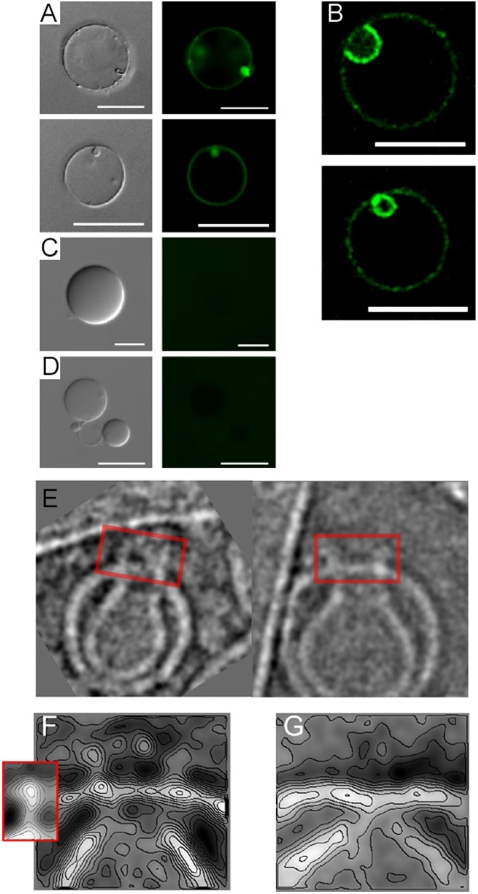
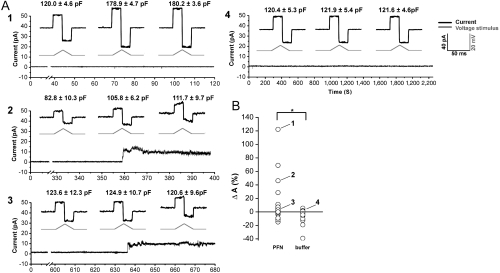
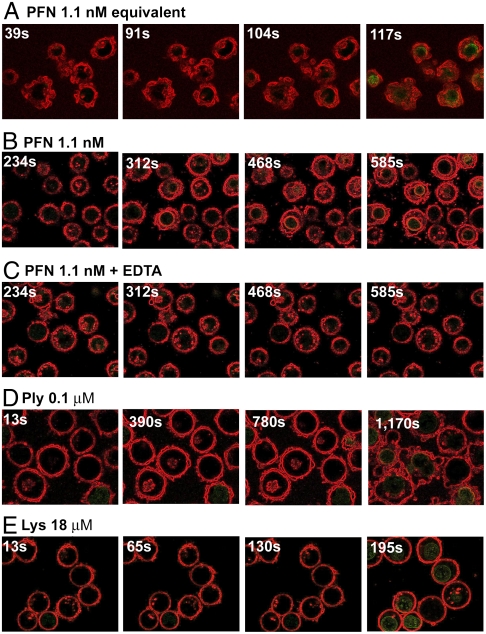
The cytotoxic cell granule secretory pathway is essential for immune defence. How the pore-forming protein perforin (PFN) facilitates the cytosolic delivery of granule-associated proteases (granzymes) remains enigmatic. Here we show that PFN is able to induce invaginations and formation of complete internal vesicles in giant unilamellar vesicles. Formation of internal vesicles depends on native PFN and calcium and antibody labeling shows the localization of PFN at the invaginations. This vesiculation is recapitulated in large unilamellar vesicles and in this case PFN oligomers can be seen associated with the necks of the invaginations. Capacitance measurements show PFN is able to increase a planar lipid membrane surface area in the absence of pore formation, in agreement with the ability to induce invaginations. Finally, addition of PFN to Jurkat cells causes the formation of internal vesicles prior to pore formation. PFN is capable of triggering an endocytosis-like event in addition to pore formation, suggesting a new paradigm for its role in delivering apoptosis-inducing granzymes into target cells.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

References
-
- Bolitho P, Voskoboinik I, Trapani JA, Smyth MJ. Apoptosis induced by the lymphocyte effector molecule perforin. Curr Opin Immunol. 2007;19:339–347. - PubMed
-
- Voskoboinik I, Smyth MJ, Trapani JA. Perforin-mediated target-cell death and immune homeostasis. Nat Rev Immunol. 2006;6:940–952. - PubMed
-
- Metkar SS, et al. Cytotoxic cell granule-mediated apoptosis: Perforin delivers granzyme B-serglycin complexes into target cells without plasma membrane pore formation. Immunity. 2002;16:417–428. - PubMed
-
- Podack ER, Dennert G. Assembly of two types of tubules with putative cytolytic function by cloned natural killer cells. Nature. 1983;302:442–445. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
