

Asymmetry and aging of mycobacterial cells lead to variable growth and antibiotic susceptibility
- PMID: 22174129
- PMCID: PMC3397429
- DOI: 10.1126/science.1216166
Asymmetry and aging of mycobacterial cells lead to variable growth and antibiotic susceptibility
Abstract
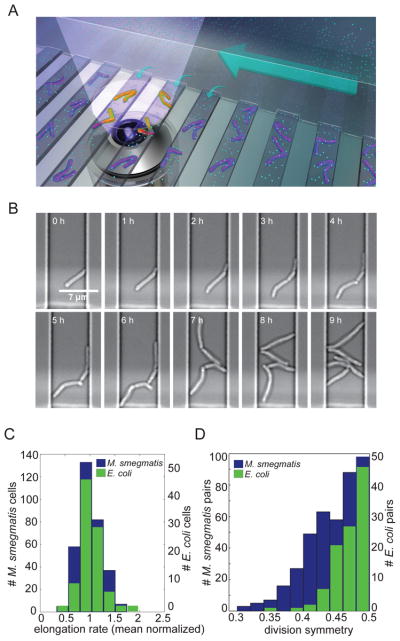
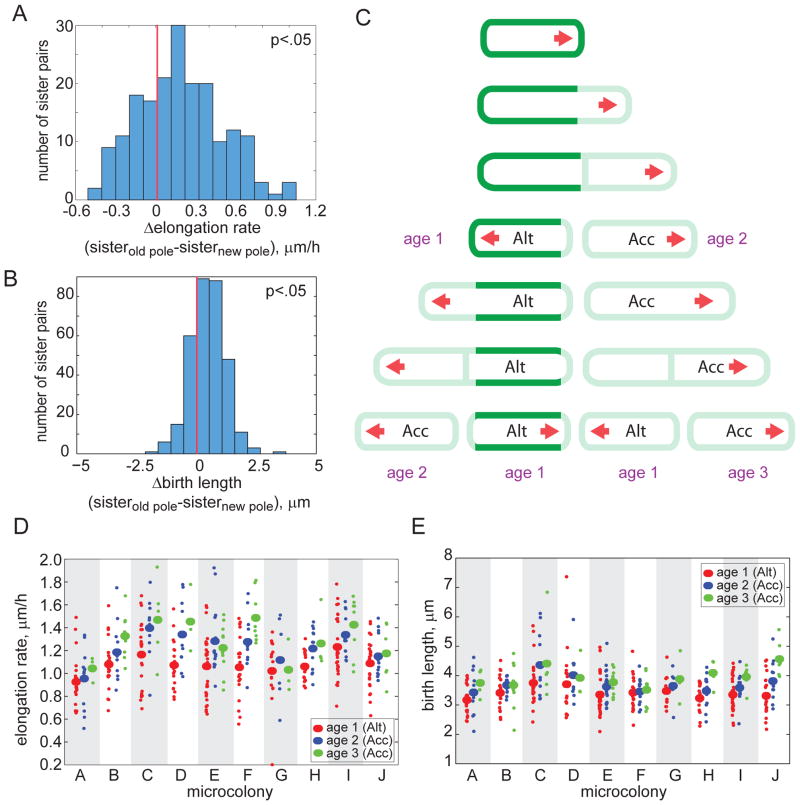
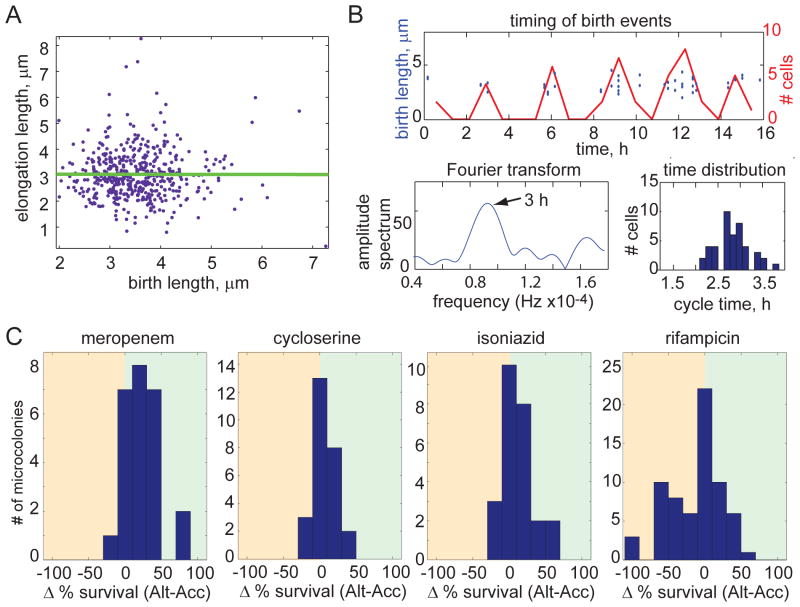
Cells use both deterministic and stochastic mechanisms to generate cell-to-cell heterogeneity, which enables the population to better withstand environmental stress. Here we show that, within a clonal population of mycobacteria, there is deterministic heterogeneity in elongation rate that arises because mycobacteria grow in an unusual, unipolar fashion. Division of the asymmetrically growing mother cell gives rise to daughter cells that differ in elongation rate and size. Because the mycobacterial cell division cycle is governed by time, not cell size, rapidly elongating cells do not divide more frequently than slowly elongating cells. The physiologically distinct subpopulations of cells that arise through asymmetric growth and division are differentially susceptible to clinically important classes of antibiotics.
Figures

Comment in
-
Asymmetric cell division: a persistent issue?Dev Cell. 2012 Feb 14;22(2):235-6. doi: 10.1016/j.devcel.2012.01.016. Dev Cell. 2012. PMID: 22340488 Free PMC article.
References
-
- Cohn DL, Catlin BJ, Peterson KL, Judson FN, Sbarbaro JA. A 62-dose, 6-month therapy for pulmonary and extrapulmonary tuberculosis. A twice-weekly, directly observed, and cost-effective regimen. Ann Intern Med. 1990;112:407–415. - PubMed
-
- Elzinga G, Raviglione MC, Maher D. Scale up: meeting targets in global tuberculosis control. Lancet. 2004;363:814–819. - PubMed
-
-
Materials and methods are available as supporting material on Science online.
-
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
