

A receptor-based switch that regulates anthrax toxin pore formation
- PMID: 22174672
- PMCID: PMC3234216
- DOI: 10.1371/journal.ppat.1002354
A receptor-based switch that regulates anthrax toxin pore formation
Abstract
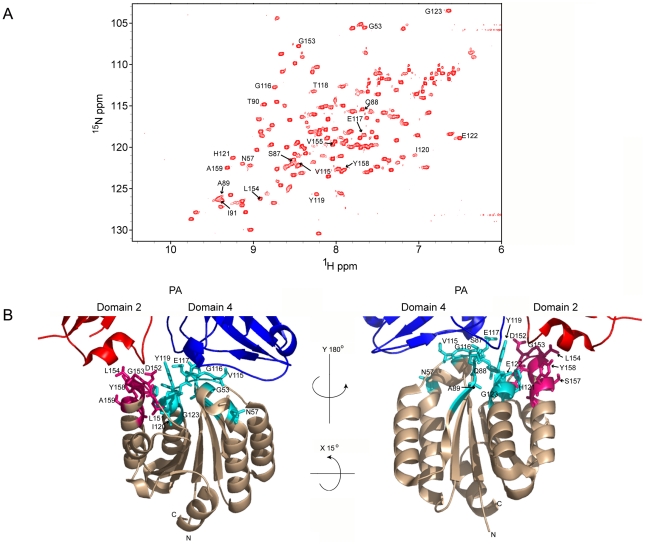
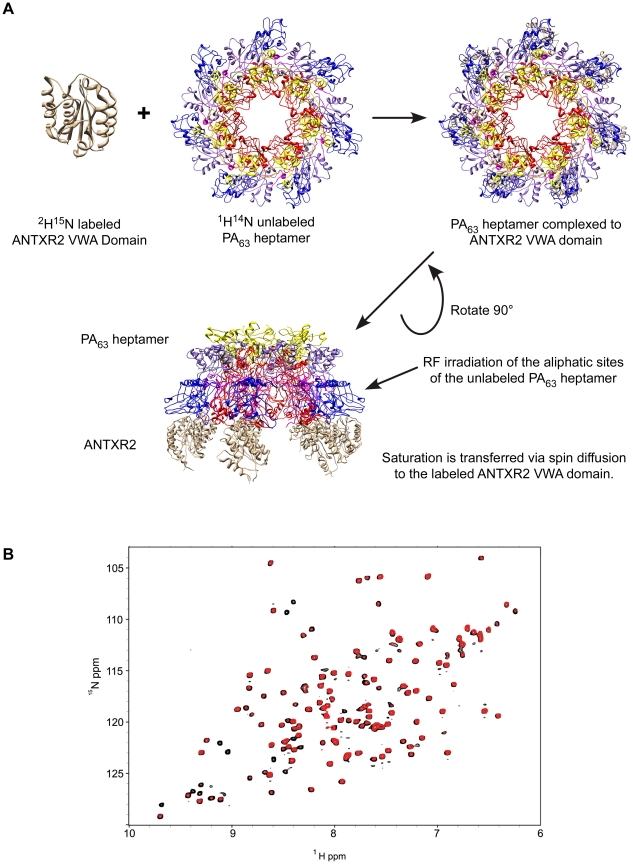
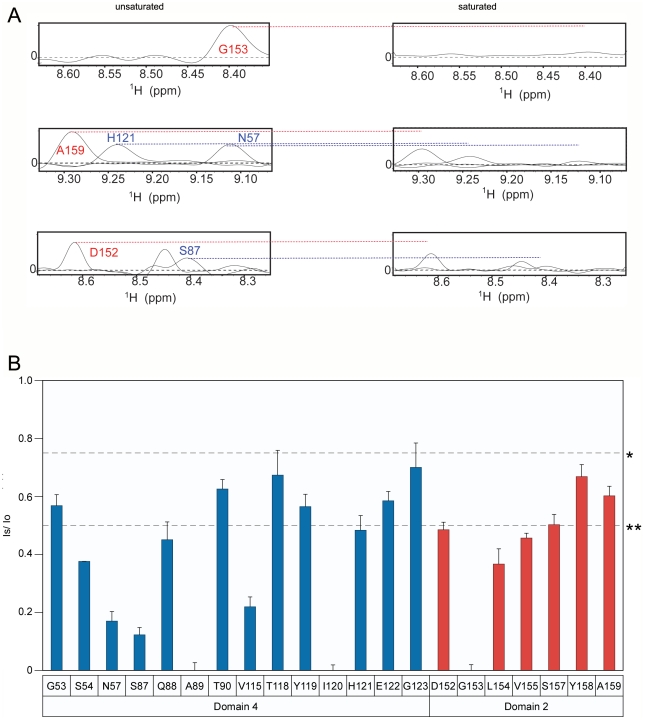
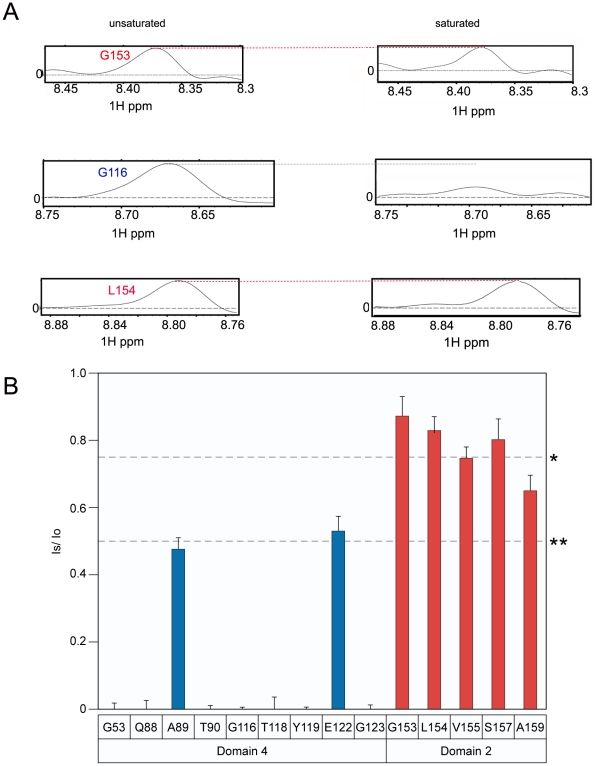
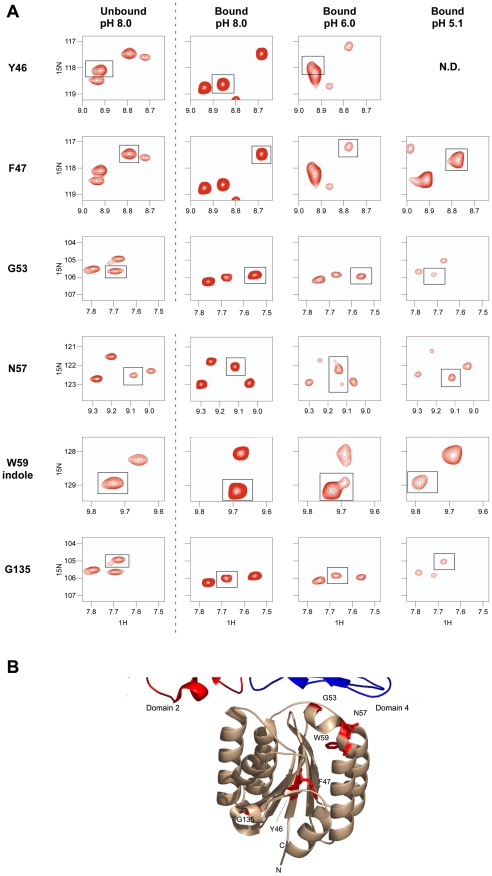
Cellular receptors can act as molecular switches, regulating the sensitivity of microbial proteins to conformational changes that promote cellular entry. The activities of these receptor-based switches are only partially understood. In this paper, we sought to understand the mechanism that underlies the activity of the ANTXR2 anthrax toxin receptor-based switch that binds to domains 2 and 4 of the protective antigen (PA) toxin subunit. Receptor-binding restricts structural changes within the heptameric PA prepore that are required for pore conversion to an acidic endosomal compartment. The transfer cross-saturation (TCS) NMR approach was used to monitor changes in the heptameric PA-receptor contacts at different steps during prepore-to-pore conversion. These studies demonstrated that receptor contact with PA domain 2 is weakened prior to pore conversion, defining a novel intermediate in this pathway. Importantly, ANTXR2 remained bound to PA domain 4 following pore conversion, suggesting that the bound receptor might influence the structure and/or function of the newly formed pore. These studies provide new insights into the function of a receptor-based molecular switch that controls anthrax toxin entry into cells.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
The Disulfide Bond Cys255-Cys279 in the Immunoglobulin-Like Domain of Anthrax Toxin Receptor 2 Is Required for Membrane Insertion of Anthrax Protective Antigen Pore.PLoS One. 2015 Jun 24;10(6):e0130832. doi: 10.1371/journal.pone.0130832. eCollection 2015. PLoS One. 2015. PMID: 26107617 Free PMC article.
-
Disulfide bonds in the ectodomain of anthrax toxin receptor 2 are required for the receptor-bound protective-antigen pore to function.PLoS One. 2010 May 10;5(5):e10553. doi: 10.1371/journal.pone.0010553. PLoS One. 2010. PMID: 20479891 Free PMC article.
-
Structure of heptameric protective antigen bound to an anthrax toxin receptor: a role for receptor in pH-dependent pore formation.Proc Natl Acad Sci U S A. 2004 Sep 7;101(36):13147-51. doi: 10.1073/pnas.0405405101. Epub 2004 Aug 23. Proc Natl Acad Sci U S A. 2004. PMID: 15326297 Free PMC article.
-
Receptors of anthrax toxin and cell entry.Mol Aspects Med. 2009 Dec;30(6):406-12. doi: 10.1016/j.mam.2009.08.007. Epub 2009 Sep 2. Mol Aspects Med. 2009. PMID: 19732789 Free PMC article. Review.
-
Interactions between anthrax toxin receptors and protective antigen.Curr Opin Microbiol. 2005 Feb;8(1):106-12. doi: 10.1016/j.mib.2004.12.005. Curr Opin Microbiol. 2005. PMID: 15694864 Review.
Cited by
-
Effect of late endosomal DOBMP lipid and traditional model lipids of electrophysiology on the anthrax toxin channel activity.Biochim Biophys Acta Biomembr. 2018 Nov;1860(11):2192-2203. doi: 10.1016/j.bbamem.2018.08.011. Epub 2018 Aug 23. Biochim Biophys Acta Biomembr. 2018. PMID: 30409515 Free PMC article.
-
Bacillus anthracis edema factor substrate specificity: evidence for new modes of action.Toxins (Basel). 2012 Jul;4(7):505-35. doi: 10.3390/toxins4070505. Epub 2012 Jul 6. Toxins (Basel). 2012. PMID: 22852066 Free PMC article. Review.
-
Anthrax toxin protective antigen integrates poly-γ-D-glutamate and pH signals to sense the optimal environment for channel formation.Proc Natl Acad Sci U S A. 2012 Nov 6;109(45):18378-83. doi: 10.1073/pnas.1208280109. Epub 2012 Oct 24. Proc Natl Acad Sci U S A. 2012. PMID: 23100533 Free PMC article.
-
Human monoclonal anti-protective antigen antibody for the low-dose post-exposure prophylaxis and treatment of Anthrax.BMC Infect Dis. 2018 Dec 10;18(1):640. doi: 10.1186/s12879-018-3542-6. BMC Infect Dis. 2018. PMID: 30526504 Free PMC article.
-
Designing inhibitors of anthrax toxin.Expert Opin Drug Discov. 2014 Mar;9(3):299-318. doi: 10.1517/17460441.2014.877884. Epub 2014 Jan 22. Expert Opin Drug Discov. 2014. PMID: 24447197 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
