

IRGM is a common target of RNA viruses that subvert the autophagy network
- PMID: 22174682
- PMCID: PMC3234227
- DOI: 10.1371/journal.ppat.1002422
IRGM is a common target of RNA viruses that subvert the autophagy network
Abstract
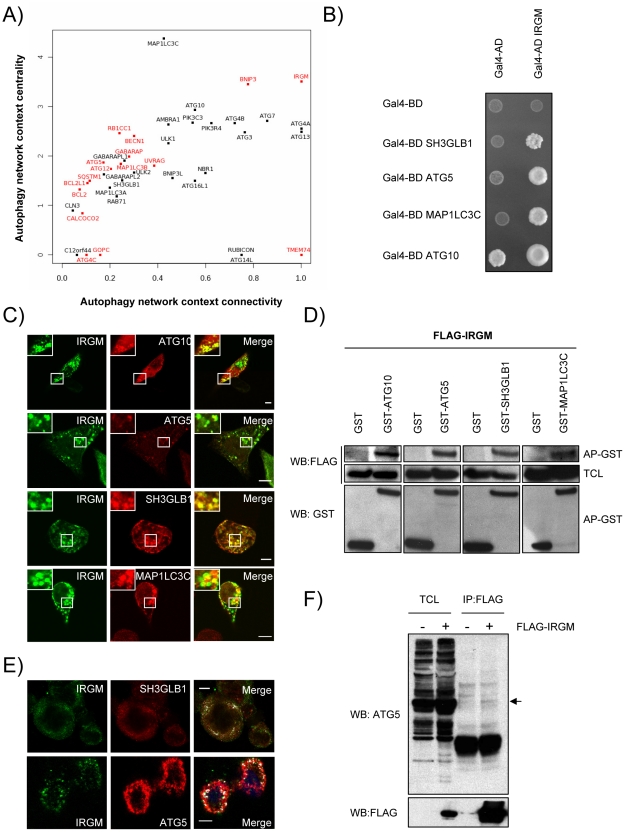
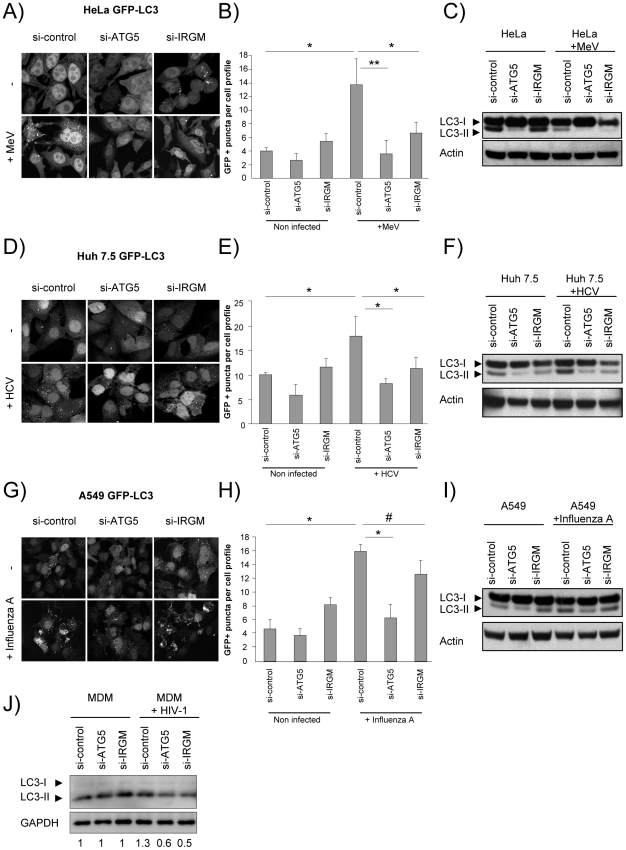
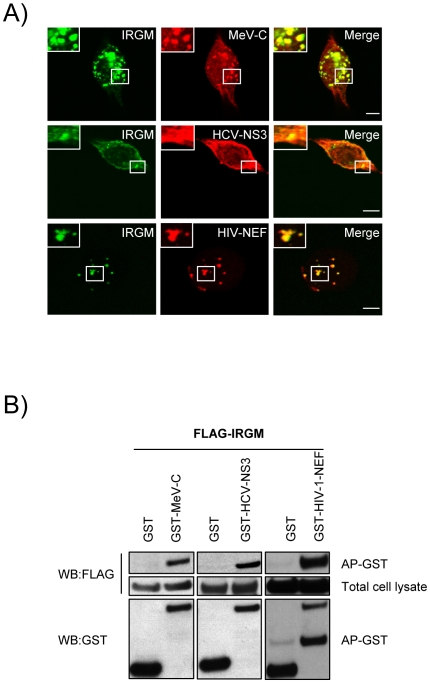
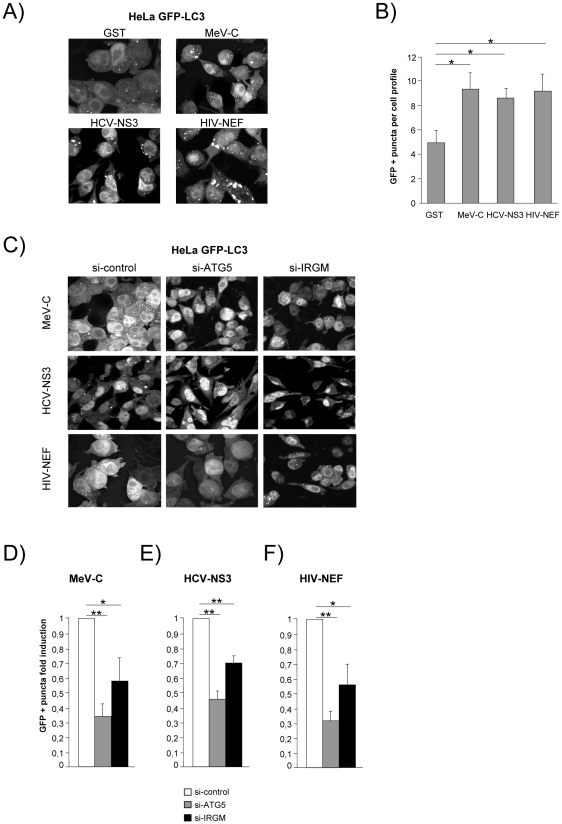
Autophagy is a conserved degradative pathway used as a host defense mechanism against intracellular pathogens. However, several viruses can evade or subvert autophagy to insure their own replication. Nevertheless, the molecular details of viral interaction with autophagy remain largely unknown. We have determined the ability of 83 proteins of several families of RNA viruses (Paramyxoviridae, Flaviviridae, Orthomyxoviridae, Retroviridae and Togaviridae), to interact with 44 human autophagy-associated proteins using yeast two-hybrid and bioinformatic analysis. We found that the autophagy network is highly targeted by RNA viruses. Although central to autophagy, targeted proteins have also a high number of connections with proteins of other cellular functions. Interestingly, immunity-associated GTPase family M (IRGM), the most targeted protein, was found to interact with the autophagy-associated proteins ATG5, ATG10, MAP1CL3C and SH3GLB1. Strikingly, reduction of IRGM expression using small interfering RNA impairs both Measles virus (MeV), Hepatitis C virus (HCV) and human immunodeficiency virus-1 (HIV-1)-induced autophagy and viral particle production. Moreover we found that the expression of IRGM-interacting MeV-C, HCV-NS3 or HIV-NEF proteins per se is sufficient to induce autophagy, through an IRGM dependent pathway. Our work reveals an unexpected role of IRGM in virus-induced autophagy and suggests that several different families of RNA viruses may use common strategies to manipulate autophagy to improve viral infectivity.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures


Comment in
-
Autophagy and RNA virus interactomes reveal IRGM as a common target.Autophagy. 2012 Jul 1;8(7):1136-7. doi: 10.4161/auto.20339. Epub 2012 Jun 22. Autophagy. 2012. PMID: 22722598 Free PMC article.
References
-
- Xie Z, Klionsky DJ. Autophagosome formation: core machinery and adaptations. Nat Cell Biol. 2007;9:1102–1109. - PubMed
-
- Levine B, Klionsky DJ. Development by self-digestion: molecular mechanisms and biological functions of autophagy. Dev Cell. 2004;6:463–477. - PubMed
-
- Furuya N, Yu J, Byfield M, Pattingre S, Levine B. The evolutionarily conserved domain of Beclin 1 is required for Vps34 binding, autophagy and tumor suppressor function. Autophagy. 2005;1:46–52. - PubMed
-
- Zeng X, Overmeyer JH, Maltese WA. Functional specificity of the mammalian Beclin-Vps34 PI 3-kinase complex in macroautophagy versus endocytosis and lysosomal enzyme trafficking. J Cell Sci. 2006;119:259–270. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
