

Small heat shock proteins and α-crystallins: dynamic proteins with flexible functions
- PMID: 22177323
- PMCID: PMC3460807
- DOI: 10.1016/j.tibs.2011.11.005
Small heat shock proteins and α-crystallins: dynamic proteins with flexible functions
Abstract
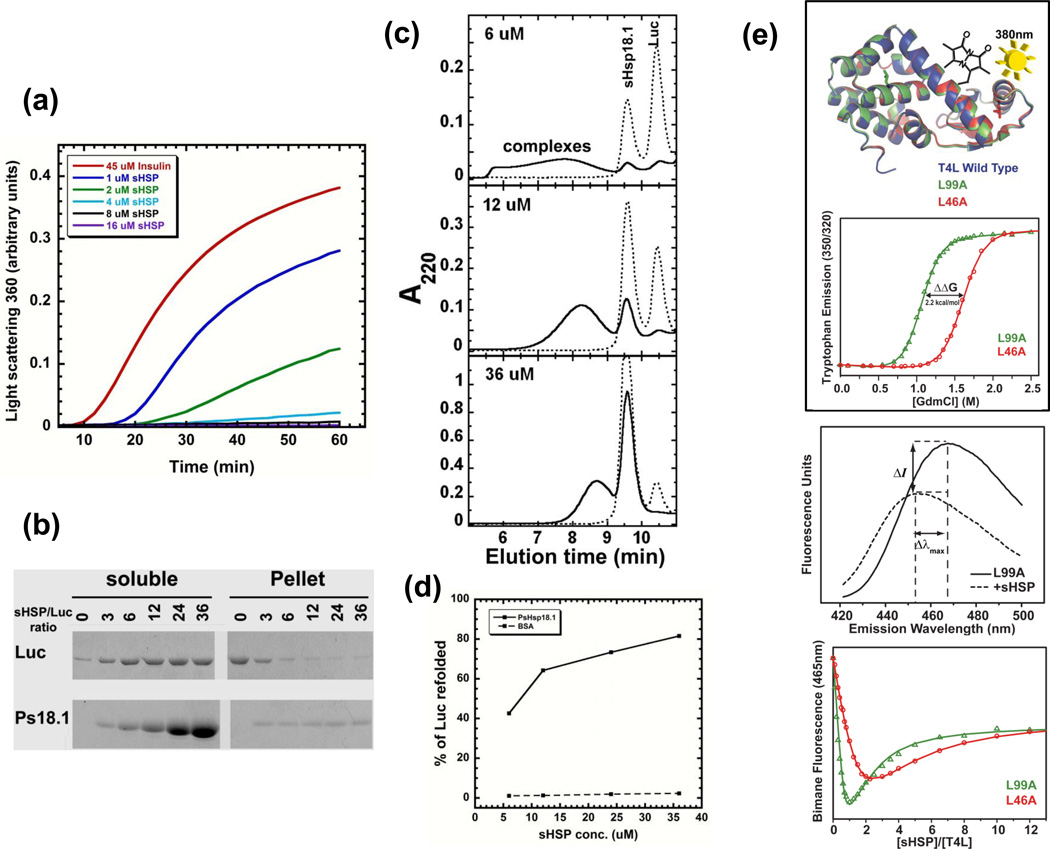
The small heat shock proteins (sHSPs) and the related α-crystallins (αCs) are virtually ubiquitous proteins that are strongly induced by a variety of stresses, but that also function constitutively in multiple cell types in many organisms. Extensive research has demonstrated that a majority of sHSPs and αCs can act as ATP-independent molecular chaperones by binding denaturing proteins and thereby protecting cells from damage due to irreversible protein aggregation. As a result of their diverse evolutionary history, their connection to inherited human diseases, and their novel protein dynamics, sHSPs and αCs are of significant interest to many areas of biology and biochemistry. However, it is increasingly clear that no single model is sufficient to describe the structure, function or mechanism of action of sHSPs and αCs. In this review, we discuss recent data that provide insight into the variety of structures of these proteins, their dynamic behavior, how they recognize substrates, and their many possible cellular roles.
Copyright © 2011 Elsevier Ltd. All rights reserved.
Figures






References
-
- Tyedmers J, et al. Cellular strategies for controlling protein aggregation. Nature Rev.Mol. Cell Biol. 2010;11:777–788. - PubMed
-
- Hartl FU, et al. Molecular chaperones in protein folding and proteostasis. Nature. 2011;475:324–332. - PubMed
-
- Jakob U, et al. Small heat shock proteins are molecular chaperones. J. Biol. Chem. 1993;268:1517–1520. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
