

Sensing sound: molecules that orchestrate mechanotransduction by hair cells
- PMID: 22177415
- PMCID: PMC3310959
- DOI: 10.1016/j.tins.2011.10.007
Sensing sound: molecules that orchestrate mechanotransduction by hair cells
Abstract
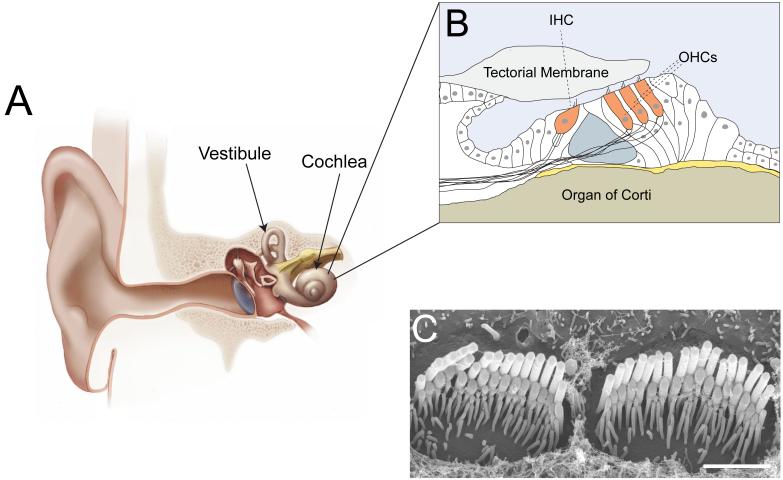
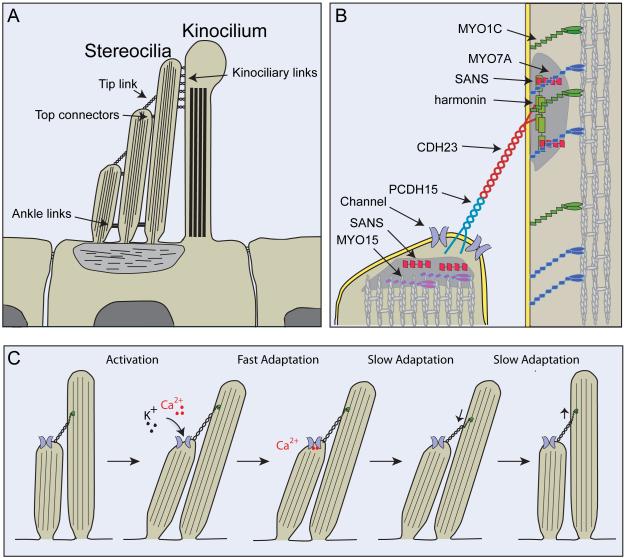
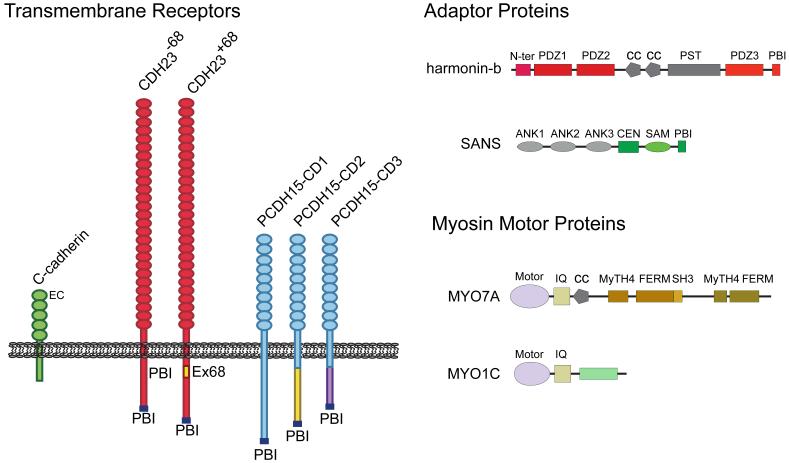
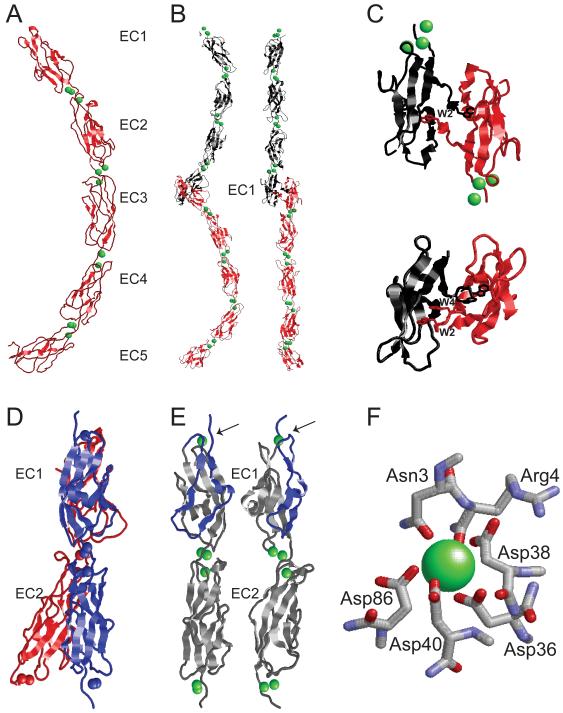
Animals use acoustic signals to communicate and to obtain information about their environment. The processing of acoustic signals is initiated at auditory sense organs, where mechanosensory hair cells convert sound-induced vibrations into electrical signals. Although the biophysical principles underlying the mechanotransduction process in hair cells have been characterized in much detail over the past 30 years, the molecular building-blocks of the mechanotransduction machinery have proved to be difficult to determine. We review here recent studies that have both identified some of these molecules and established the mechanisms by which they regulate the activity of the still-elusive mechanotransduction channel.
Copyright © 2011 Elsevier Ltd. All rights reserved.
Figures
Similar articles
-
Molecular Identity of the Mechanotransduction Channel in Hair Cells: Not Quiet There Yet.J Neurosci. 2016 Oct 26;36(43):10927-10934. doi: 10.1523/JNEUROSCI.1149-16.2016. J Neurosci. 2016. PMID: 27798175 Free PMC article.
-
The elusive mechanotransduction machinery of hair cells.Curr Opin Neurobiol. 2015 Oct;34:172-9. doi: 10.1016/j.conb.2015.08.006. Epub 2015 Sep 3. Curr Opin Neurobiol. 2015. PMID: 26342686 Free PMC article. Review.
-
Mechanisms in cochlear hair cell mechano-electrical transduction for acquisition of sound frequency and intensity.Cell Mol Life Sci. 2021 Jun;78(12):5083-5094. doi: 10.1007/s00018-021-03840-8. Epub 2021 Apr 19. Cell Mol Life Sci. 2021. PMID: 33871677 Free PMC article. Review.
-
The mechanotransduction machinery of hair cells.Sci Signal. 2009 Aug 25;2(85):pt5. doi: 10.1126/scisignal.285pt5. Sci Signal. 2009. PMID: 19706872
-
Modeling auditory transducer dynamics.Curr Opin Otolaryngol Head Neck Surg. 2009 Oct;17(5):400-6. doi: 10.1097/MOO.0b013e3283303443. Curr Opin Otolaryngol Head Neck Surg. 2009. PMID: 19625966 Review.
Cited by
-
The proteome of mouse vestibular hair bundles over development.Sci Data. 2015 Sep 15;2:150047. doi: 10.1038/sdata.2015.47. eCollection 2015. Sci Data. 2015. PMID: 26401315 Free PMC article.
-
Deletion of Kcnj16 in Mice Does Not Alter Auditory Function.Front Cell Dev Biol. 2021 Feb 22;9:630361. doi: 10.3389/fcell.2021.630361. eCollection 2021. Front Cell Dev Biol. 2021. PMID: 33693002 Free PMC article.
-
Recent Advances in Artificial Sensory Neurons: Biological Fundamentals, Devices, Applications, and Challenges.Nanomicro Lett. 2024 Nov 13;17(1):61. doi: 10.1007/s40820-024-01550-x. Nanomicro Lett. 2024. PMID: 39537845 Free PMC article. Review.
-
Lis1 mediates planar polarity of auditory hair cells through regulation of microtubule organization.Development. 2013 Apr;140(8):1785-95. doi: 10.1242/dev.089763. Development. 2013. PMID: 23533177 Free PMC article.
-
Developmental changes in the cochlear hair cell mechanotransducer channel and their regulation by transmembrane channel-like proteins.J Gen Physiol. 2013 Jan;141(1):141-8. doi: 10.1085/jgp.201210913. J Gen Physiol. 2013. PMID: 23277480 Free PMC article.
References
-
- Meyer AC, Moser T. Structure and function of cochlear afferent innervation. Curr Opin Otolaryngol Head Neck Surg. 2010;18:441–446. - PubMed
-
- Richardson GP, et al. How the genetics of deafness illuminates auditory physiology. Annual review of physiology. 2011;73:311–334. - PubMed
-
- Pickles JO, et al. Cross-links between stereocilia in the guinea pig organ of Corti, and their possible relation to sensory transduction. Hear Res. 1984;15:103–112. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
