

Ser/ Thr residues at α3/β5 loop of Gαs are important in morphine-induced adenylyl cyclase sensitization but not mitogen-activated protein kinase phosphorylation
- PMID: 22177524
- PMCID: PMC4540596
- DOI: 10.1111/j.1742-4658.2011.08459.x
Ser/ Thr residues at α3/β5 loop of Gαs are important in morphine-induced adenylyl cyclase sensitization but not mitogen-activated protein kinase phosphorylation
Abstract
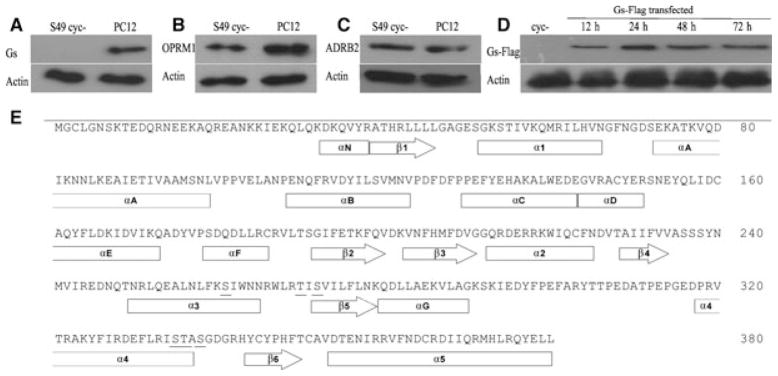
The signaling switch of β2-adrenergic and μ(1) -opioid receptors from stimulatory G-protein (G(αs) ) to inhibitory G-protein (G(αi) ) (and vice versa) influences adenylyl cyclase (AC) and extracellular-regulated kinase (ERK)1/2 activation. Post-translational modifications, including dephosphorylation of G(αs) , enhance opioid receptor coupling to G(αs) . In the present study, we substituted the Ser/Thr residues of G(αs) at the α3/β5 and α4/β6 loops aiming to study the role of G(αs) lacking Ser/Thr phosphorylation with respect to AC sensitization and mitogen-activated protein kinase activation. Isoproterenol increased the cAMP concentration (EC(50) = 22.8 ± 3.4 μm) in G(αs) -transfected S49 cyc- cells but not in nontransfected cells. However, there was no significant difference between the G(αs) -wild-type (wt) and mutants. Morphine (10 μm) inhibited AC activity more efficiently in cyc- compared to G(αs) -wt introduced cells (P < 0.05); however, we did not find a notable difference between G(αs) -wt and mutants. Interestingly, G(αs) -wt transfected cells showed more sensitization with respect to AC after chronic morphine compared to nontransfected cells (101 ± 12% versus 34 ± 6%; P < 0.001); μ1-opioid receptor interacted with G(αs) , and both co-immunoprecipitated after chronic morphine exposure. Furthermore, mutation of T270A and S272A (P < 0.01), as well as T270A, S272A and S261A (P < 0.05), in α3/β5, resulted in a higher level of AC supersensitization. ERK1/2 phosphorylation was rapidly induced by isoproterenol (by 9.5 ± 2.4-fold) and morphine (22 ± 2.2-fold) in G(αs) -transfected cells; mutations of α3/β5 and α4/β6 did not affect the pattern or extent of mitogen-activated protein kinase activation. The findings of the present study show that G(αs) interacts with the μ1-opioid receptor, and the Ser/Thr mutation to Ala at the α3/β5 loop of G(αs) enhances morphine-induced AC sensitization. In addition, G(αs) was required for the rapid phosphorylation of ERK1/2 by isoproterenol but not morphine.
© 2011 The Authors Journal compilation © 2011 FEBS.
Figures





Similar articles
-
Protein kinase A-mediated phosphorylation of the beta 2-adrenergic receptor regulates its coupling to Gs and Gi. Demonstration in a reconstituted system.J Biol Chem. 2002 Aug 23;277(34):31249-56. doi: 10.1074/jbc.M202753200. Epub 2002 Jun 12. J Biol Chem. 2002. PMID: 12063255
-
Endogenous RGS protein action modulates mu-opioid signaling through Galphao. Effects on adenylyl cyclase, extracellular signal-regulated kinases, and intracellular calcium pathways.J Biol Chem. 2003 Mar 14;278(11):9418-25. doi: 10.1074/jbc.M208885200. Epub 2003 Jan 10. J Biol Chem. 2003. PMID: 12524446
-
Regulation of adenylyl cyclase, ERK1/2, and CREB by Gz following acute and chronic activation of the delta-opioid receptor.J Neurochem. 2000 Apr;74(4):1685-93. doi: 10.1046/j.1471-4159.2000.0741685.x. J Neurochem. 2000. PMID: 10737627
-
Hydrogen sulfide attenuates opioid dependence by suppression of adenylate cyclase/cAMP pathway.Antioxid Redox Signal. 2014 Jan 1;20(1):31-41. doi: 10.1089/ars.2012.5119. Epub 2013 Jul 11. Antioxid Redox Signal. 2014. PMID: 23682813 Free PMC article.
-
Activation of a beta 2-adrenergic receptor/Gs alpha fusion protein elicits a desensitization-resistant cAMP signal capable of inhibiting proliferation of two cancer cell lines.Recept Channels. 1997;5(1):41-51. Recept Channels. 1997. PMID: 9272575
Cited by
-
The Neuroprotective Effect of Lithium in cannabinoid Dependence is Mediated through Modulation of Cyclic AMP, ERK1/2 and GSK-3β Phosphorylation in Cerebellar Granular Neurons of Rat.Iran J Pharm Res. 2015 Fall;14(4):1123-35. Iran J Pharm Res. 2015. PMID: 26664379 Free PMC article.
-
Receptor-Arrestin Interactions: The GPCR Perspective.Biomolecules. 2021 Feb 4;11(2):218. doi: 10.3390/biom11020218. Biomolecules. 2021. PMID: 33557162 Free PMC article. Review.
References
-
- Cabrera-Vera TM, Vanhauwe J, Thomas TO, Medkova M, Preininger A, Mazzoni MR, Hamm HE. Insights into G protein structure, function, and regulation. Endocr Rev. 2003;24:765–781. - PubMed
-
- Muramatsu T, Suwa M. Statistical analysis and prediction of functional residues effective for GPCR-G-protein coupling selectivity. Protein Eng Des Sel. 2006;19:277–283. - PubMed
-
- Slessareva JE, Ma H, Depree KM, Flood LA, Bae H, Cabrera-Vera TM, Hamm HE, Graber SG. Closely related G-protein-coupled receptors use multiple and distinct domains on G-protein alpha-subunits for selective coupling. J Biol Chem. 2003;278:50530–50536. - PubMed
-
- Heydorn A, Ward RJ, Jorgensen R, Rosenkilde MM, Frimurer TM, Milligan G, Kostenis E. Identification of a novel site within G protein alpha subunits important for specificity of receptor-G protein interaction. Mol Pharmacol. 2004;66:250–259. - PubMed
-
- Sunahara RK, Tesmer JJ, Gilman AG, Sprang SR. Crystal structure of the adenylyl cyclase activator Gsalpha. Science. 1997;278:1943–1947. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
