

Review
doi: 10.1016/j.conb.2011.11.013.
Epub 2011 Dec 15.
Sniffing and whisking in rodents
Affiliations
- PMID: 22177596
- PMCID: PMC4934665
- DOI: 10.1016/j.conb.2011.11.013
Item in Clipboard
Review
Sniffing and whisking in rodents
Curr Opin Neurobiol.
2012 Apr.
Abstract
Sniffing and whisking are two rhythmic orofacial motor activities that enable rodents to localize and track objects in their environment. They have related temporal dynamics, possibly as a result of both shared musculature and shared sensory tasks. Sniffing and whisking also constitute the overt expression of an animal's anticipation of a reward. Yet, the neuronal mechanisms that underlie the control of these behaviors have not been established. Here, we review the similarities between sniffing and whisking and suggest that such similarities indicate a mechanistic link between these two rhythmic exploratory behaviors.
Copyright © 2011 Elsevier Ltd. All rights reserved.
Figures
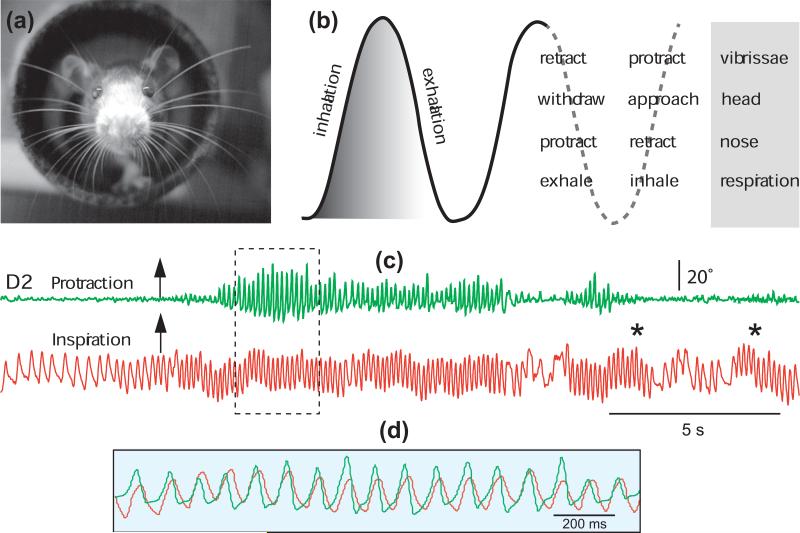
(a) Photograph of a rat whisking at the exit of a tunnel. Image was obtained in the dark under near infrared illumination. (b) Sniff and whisk cycles are coordinated with nose and head movements. Adapted from Welker [2]. (c) Whisking occurs in phase with sniffing, but sniffing can occur without whisking; asterisks denote sniffing bouts without whisking. Overlapping traces in (d) show the framed area in (c). The motion of vibrissa D2 was monitored by high-speed videography (250 frames per second), and sniffing was recorded by means of a thermocouple implanted in the nasal cavity (unpublished data).
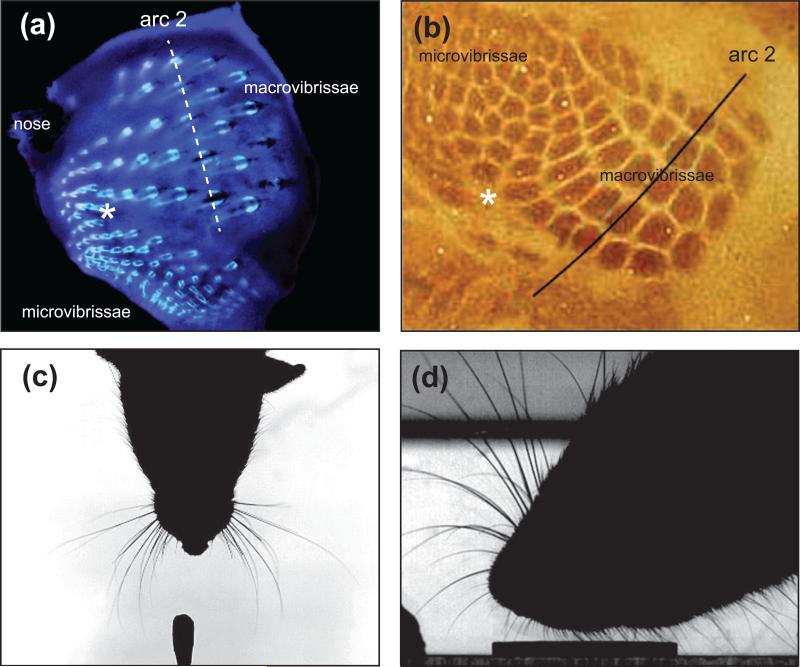
(a) Vibrissa follicles on the mystacial pad and perioral region. (b) The folliciles are mapped in a one-to-one manner in layer 4 of the primary somatosensory cortex. Note the presence of small-sized follicles, i.e., microvibrissae, that are represented by smaller sized barrels in cortex. Asterisks indicate the transition point between macro- and microvibrissae and their representation in cortex. (c) Photograph of a rat whisking upon presentation of a new odor on the cotton swab in the lower part of the image (unpublished data). (d) Photograph of a rat sampling a coin with its microvibrissae. Adapted from Fox et al. [50].
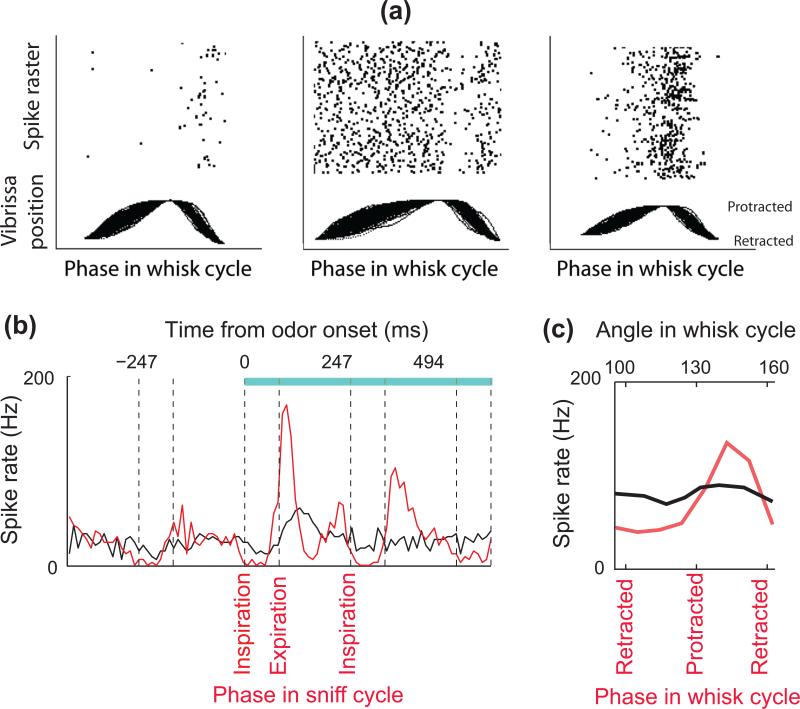
(a) Whisking solely in air modulates the activity of trigeminal ganglion cells. Unit recordings were obtained from a head-fixed rat together with simultaneous optical tracking of vibrissa position. Vibrissa movements are aligned to protraction onset, retraction onset, and end. Raster plots the spike times above the movements. Adapted from Khatri et al. [19]. (b) Peristimulus time histograms for a mitral/tufted cell in response to an odor stimulus (blue bar, stimulus duration): synchronized by odor onset (black), and temporally warped to the phase in the sniff cycle (red). Note that mitral cell activity is modulated by sniffing before the odor presentation, and that odor-induced responses are more tightly time-locked to the sniff phase than to the time after odor onset. Vertical dashed lines indicate the beginning and end of inhalation intervals. Adapted from Shusterman et al. [21]. (c) Touch response in vibrissa sensory cortex is strongly modulated by the phase in the whisk cycle. The red trace shows the spike rate of a neuron in response to object contact parsed according to the phase in the whisk cycle at which the contact occurred. The black line is the same data parsed according to the angular position of the vibrissa where the contact occurred. Unlike the case for phase, there is no significant tuning for angle. Adapted from Curtis and Kleinfeld [24].
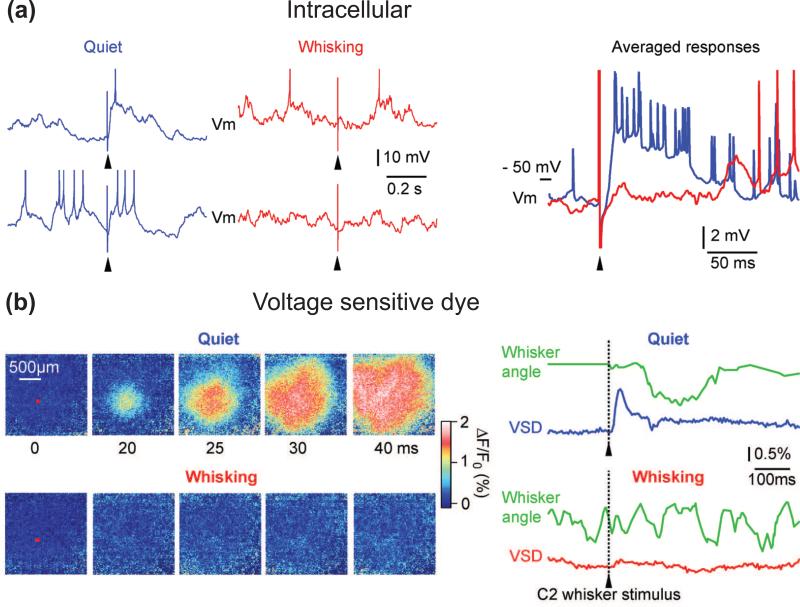
(a) Vibrissa stimuli (arrowheads) were delivered randomly during recording from a layer 4 barrel neuron. Intracellular recordings show that the depolarizing sensory response is strongly reduced during active whisking (red) compared to during quiescent wakefulness (blue). Adapted from Crochet and Petersen [23]. (b) The above state-dependent reduction in sensory processing can be imaged with voltage-sensitive dye. Passively evoked sensory responses during quiescent wakefulness have large amplitude and spread across large cortical areas, whereas the response is almost suppressed during whisking. The red square on the images at 0 ms indicates the region of interest centered on the C2 cortical column from which voltage-sensitive dye fluorescence changes are quantified in the adjacent traces (lower right). Adapted from Ferezou et al. [28].
References
-
- Welker WI. Analysis of sniffing of the albino rat. Behavior. 1964;22:223–244.
-
- Woolsey TA, Van der Loos H. The structural organization of layer IV in the somatosensory region (SI) of mouse cerebral cortex. The description of a cortical field composed of discrete cytoarchitectonic units. Brain Res. 1970;17:205–242. - PubMed
-
- Sherrey JH, Megirian D. State dependence of upper airway respiratory motoneurons: functions of the cricothyroid and nasolabial muscles of the unanesthetized rat. EEG Clin Neurophysiol. 1977;43:218–228. - PubMed
-
- Hwang JC, St John WM. Respiratory-modulated activities of motor units of the facial nerve. Respir Physiol. 1988;73:189–200. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
