

Working memory, long-term memory, and medial temporal lobe function
- PMID: 22180053
- PMCID: PMC3246590
- DOI: 10.1101/lm.024018.111
Working memory, long-term memory, and medial temporal lobe function
Abstract
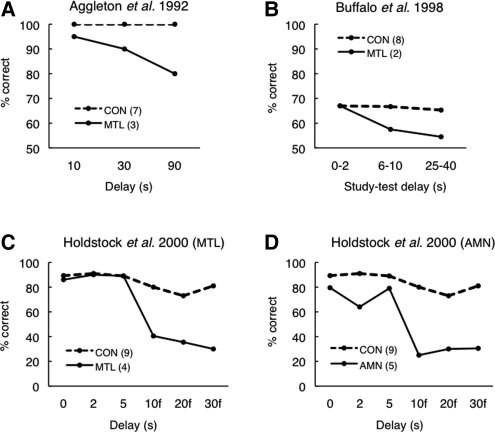
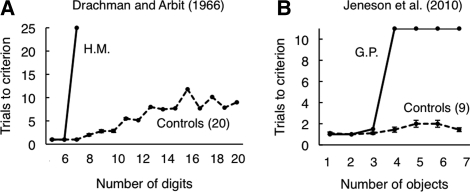
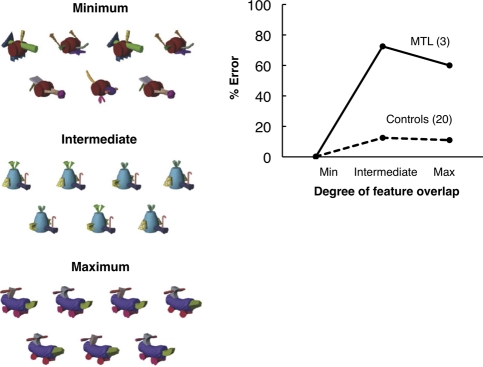
Early studies of memory-impaired patients with medial temporal lobe (MTL) damage led to the view that the hippocampus and related MTL structures are involved in the formation of long-term memory and that immediate memory and working memory are independent of these structures. This traditional idea has recently been revisited. Impaired performance in patients with MTL lesions on tasks with short retention intervals, or no retention interval, and neuroimaging findings with similar tasks have been interpreted to mean that the MTL is sometimes needed for working memory and possibly even for visual perception itself. We present a reappraisal of this interpretation. Our main conclusion is that, if the material to be learned exceeds working memory capacity, if the material is difficult to rehearse, or if attention is diverted, performance depends on long-term memory even when the retention interval is brief. This fundamental notion is better captured by the terms subspan memory and supraspan memory than by the terms short-term memory and long-term memory. We propose methods for determining when performance on short-delay tasks must depend on long-term (supraspan) memory and suggest that MTL lesions impair performance only when immediate memory and working memory are insufficient to support performance. In neuroimaging studies, MTL activity during encoding is influenced by the memory load and correlates positively with long-term retention of the material that was presented. The most parsimonious and consistent interpretation of all the data is that subspan memoranda are supported by immediate memory and working memory and are independent of the MTL.
Figures
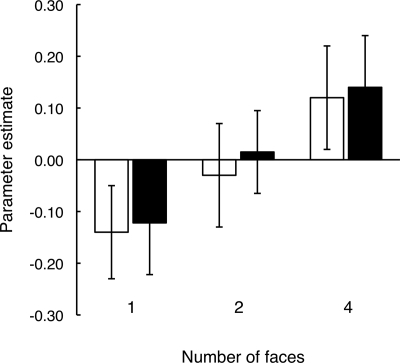
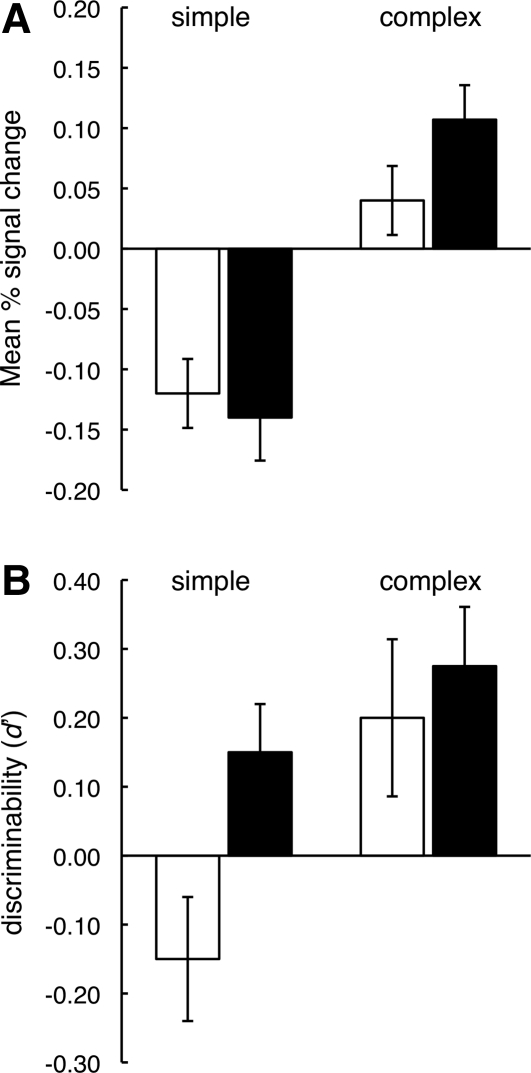
References
-
- Aggleton JP, Shaw C, Gaffan EA 1992. The performance of postencephalitic amnesic subjects on two behavioural tests of memory: Concurrent discrimination learning and delayed matching-to-sample. Cortex 28: 359–372 - PubMed
-
- Alvarez G, Cavanagh P 2004. The capacity of visual short-term memory is set both by visual information load and by number of objects. Psychol Sci 15: 106–111 - PubMed
-
- Alvarez GA, Thompson TW 2009. Overwriting and rebinding: Why feature-switch detection tasks underestimate the binding capacity of visual working memory. Vis Cogn 17: 141–159
-
- Atkinson RC, Shiffrin RM 1968. Human memory: A proposed system and its control processes. In The psychology of learning and motivation: II (ed. Spence KW, Spence JT). Academic Press, Oxford, England
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources