

Cut-like homeobox 1 and nuclear factor I/B mediate ENGRAILED2 autism spectrum disorder-associated haplotype function
- PMID: 22180456
- PMCID: PMC3298280
- DOI: 10.1093/hmg/ddr594
Cut-like homeobox 1 and nuclear factor I/B mediate ENGRAILED2 autism spectrum disorder-associated haplotype function
Abstract
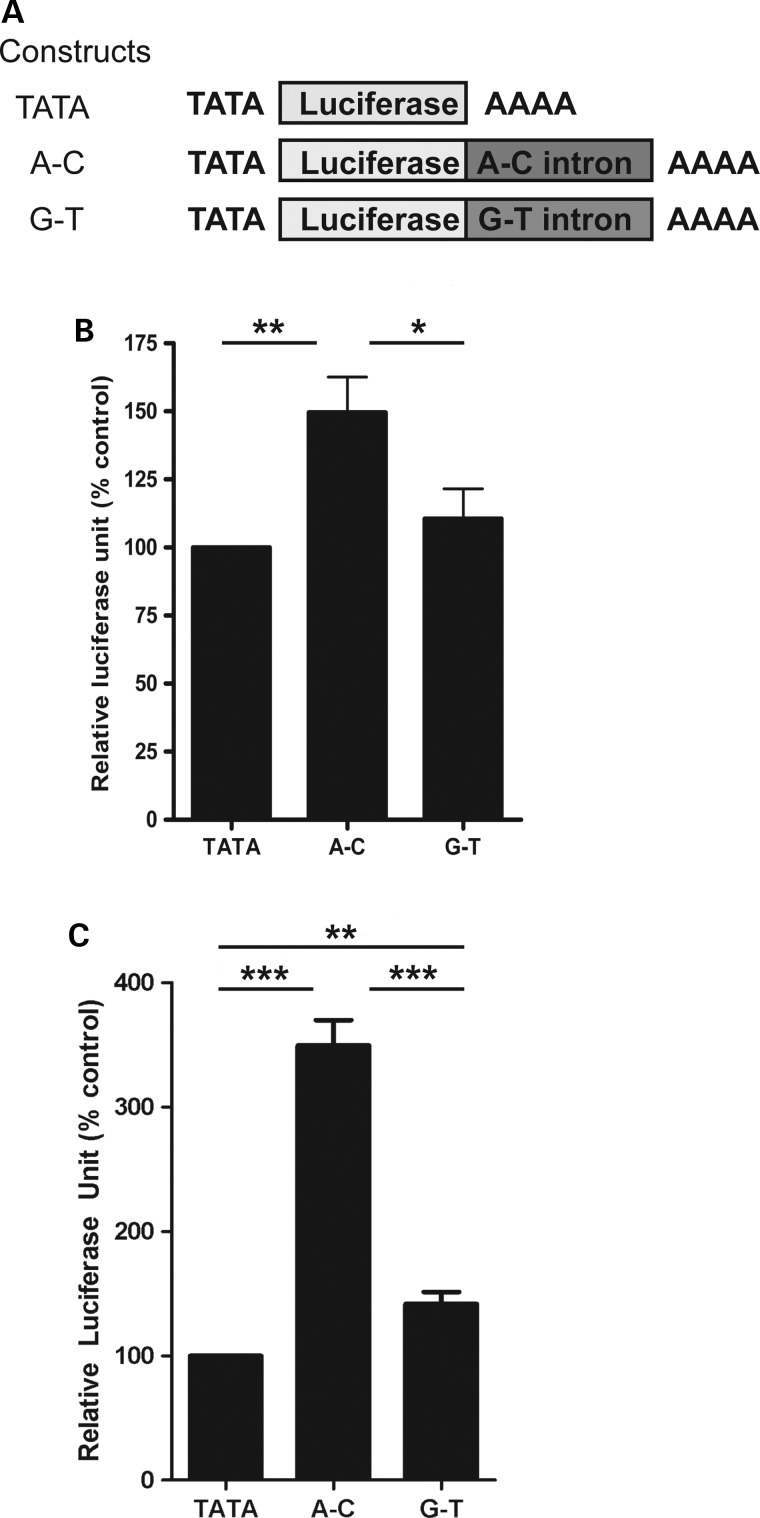
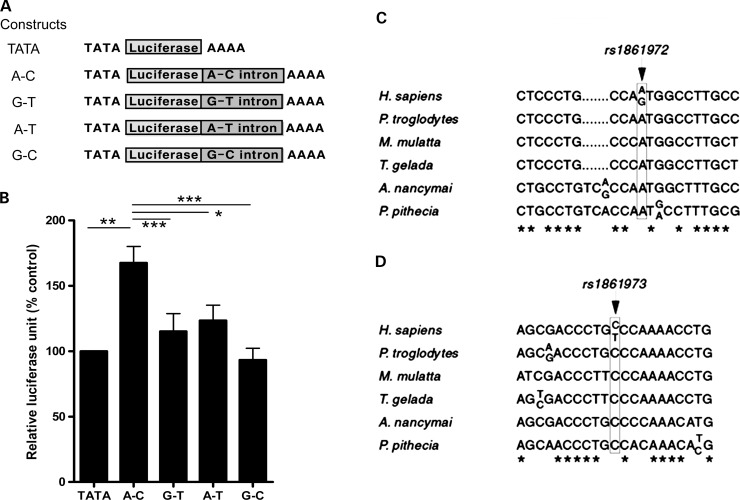
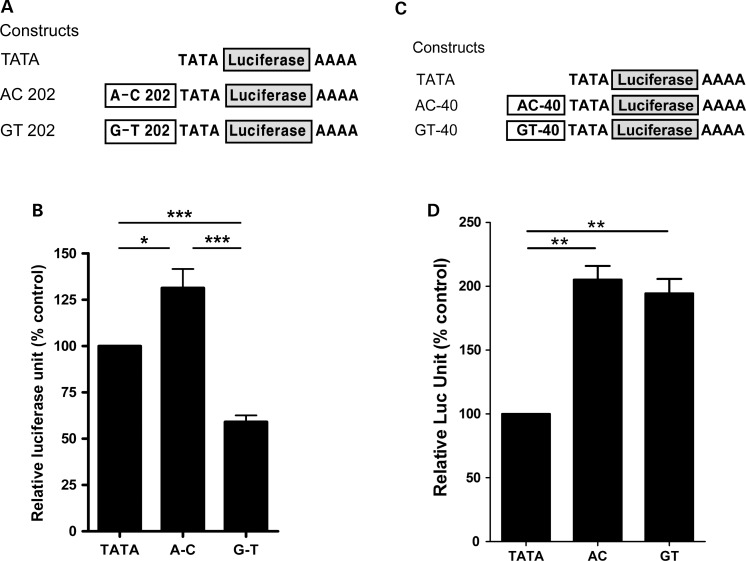
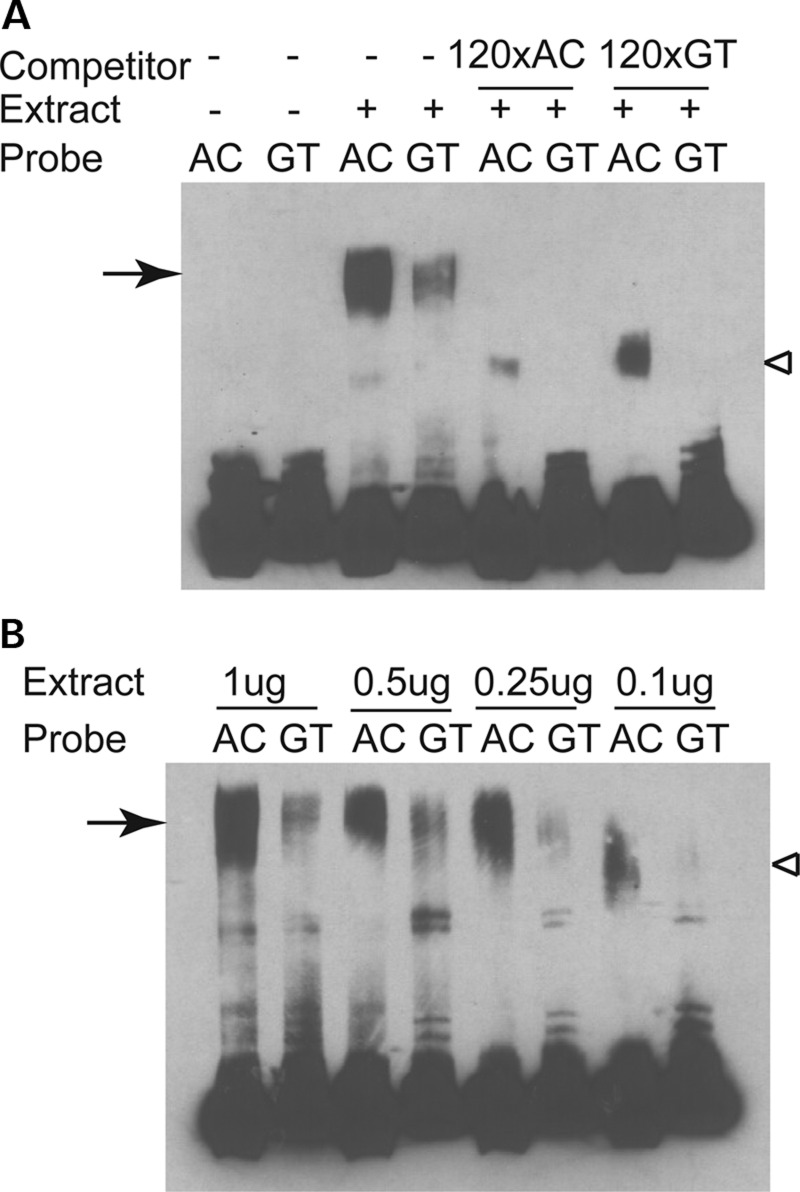
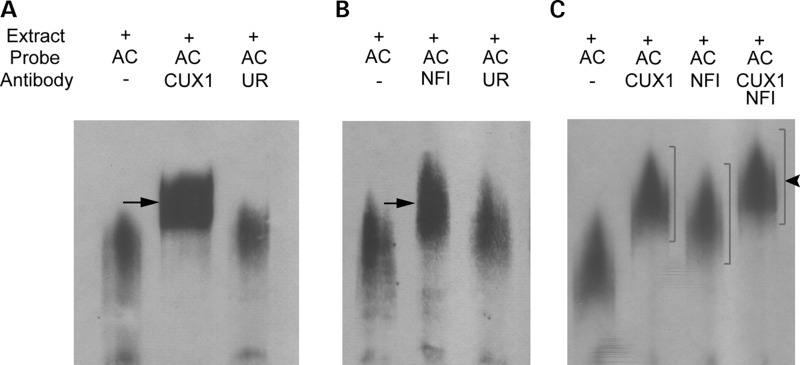
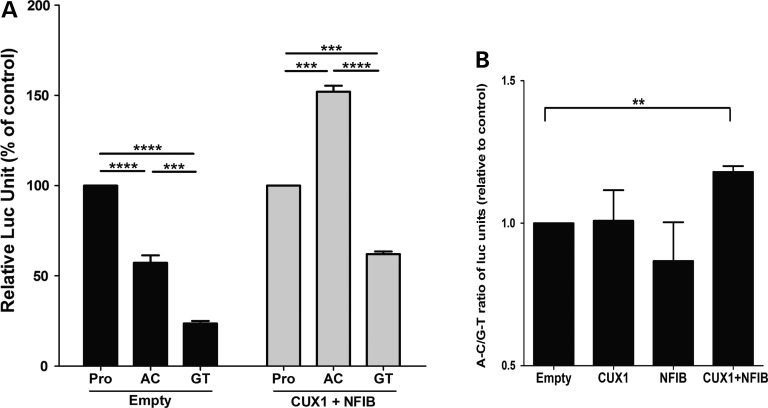
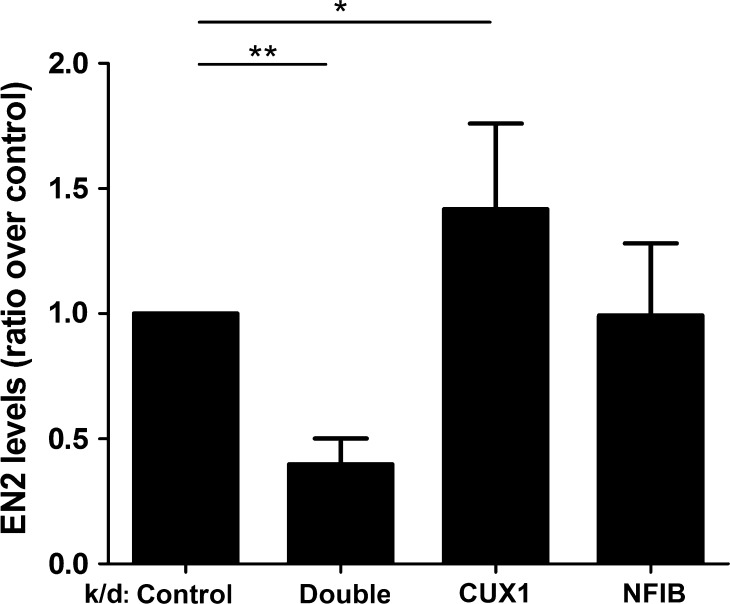
Both common and rare variants contribute to autism spectrum disorder (ASD) risk, but few variants have been established as functional. Previously we demonstrated that an intronic haplotype (rs1861972-rs1861973 A-C) in the homeobox transcription factor ENGRAILED2 (EN2) is significantly associated with ASD. Positive association has also been reported in six additional data sets, suggesting EN2 is an ASD susceptibility gene. Additional support for this possibility requires identification of functional variants that affect EN2 regulation or activity. In this study, we demonstrate that the A-C haplotype is a transcriptional activator. Luciferase (luc) assays in mouse neuronal cultures determined that the A-C haplotype increases expression levels (50%, P < 0.01, 24 h; 250%, P < 0.0001, 72 h). Mutational analysis indicates that the A-C haplotype activator function requires both associated A and C alleles. A minimal 202-bp element is sufficient for function and also specifically binds a protein complex. Mass spectrometry identified these proteins as the transcription factors, Cut-like homeobox 1 (Cux1) and nuclear factor I/B (Nfib). Subsequent antibody supershifts and chromatin immunoprecipitations demonstrated that human CUX1 and NFIB bind the A-C haplotype. Co-transfection and knock-down experiments determined that both CUX1 and NFIB are required for the A-C haplotype activator function. These data demonstrate that the ASD-associated A-C haplotype is a transcriptional activator, and both CUX1 and NFIB mediate this activity. These results provide biochemical evidence that the ASD-associated A-C haplotype is functional, further supporting EN2 as an ASD susceptibility gene.
Figures


References
-
- Dawson G. Recent advances in research on early detection, causes, biology, and treatment of autism spectrum disorders. Curr. Opin. Neurol. 2010;23:95–96. doi:10.1097/WCO.0b013e3283377644. - DOI - PubMed
-
- Freitag C.M. The genetics of autistic disorders and its clinical relevance: a review of the literature. Mol. Psychiatry. 2007;12:2–22. doi:10.1038/sj.mp.4001896. - DOI - PubMed
-
- Ronald A., Happe F., Bolton P., Butcher L.M., Price T.S., Wheelwright S., Baron-Cohen S., Plomin R. Genetic heterogeneity between the three components of the autism spectrum: a twin study. J. Am. Acad. Child Adolesc. Psychiatry. 2006;45:691–699. doi:10.1097/01.chi.0000215325.13058.9d. - DOI - PubMed
-
- Geschwind D.H. Advances in autism. Annu. Rev. Med. 2009;60:367–380. doi:10.1146/annurev.med.60.053107.121225. - DOI - PMC - PubMed
-
- State M.W. The genetics of child psychiatric disorders: focus on autism and Tourette syndrome. Neuron. 2010;68:254–269. doi:10.1016/j.neuron.2010.10.004. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
